
Method Article
Colheita de toxinas de veneno de barbeiros e outros insetos Heteropteran
Neste Artigo
Resumo
Embora muitos insetos da subordem Heteroptera (Insecta: Hemiptera) são venenosa, sua composição de veneno e as funções das suas toxinas de veneno em sua maioria são desconhecidas. Este protocolo descreve métodos para colher heteropteran venenos para mais de caracterização, utilizando a eletroestimulação, assédio e dissecação da glândula.
Resumo
Heteropteran insetos como barbeiros (Reduviidae) e bugs de água gigante (Belostomatidae) descendem de um ancestral comum de predadores e venenoso, e a maioria dos heteropterans existentes manter esta estratégia trófica. Alguns heteropterans ter transferida para alimentando-se de sangue de vertebrados (como os kissing bugs, triatomíneos; e percevejos, Cimicidae) enquanto outros foram revertidos para a alimentação das plantas (mais Pentatomomorpha). No entanto, com exceção de saliva usada por kissing bugs para facilitar a alimentação de sangue, pouco é conhecido sobre venenos de heteropteran em comparação com os venenos de aranhas, escorpiões e cobras.
Um obstáculo para a caracterização de toxinas de veneno de heteropteran é a estrutura e função das glândulas de veneno/labial, que são ambos morfologicamente complexos e executar várias funções biológicas (defesa, captura de presas e digestão extra oral). Neste artigo, descrevemos três métodos que usamos com sucesso para coletar heteropteran venenos. Primeiro, apresentamos a eletroestimulação como uma maneira conveniente para coletar o veneno que muitas vezes é letal quando injetado em animais de rapina, e que elimina a contaminação por tecido glandular. Em segundo lugar, nós mostramos que o assédio suave dos animais é suficiente para produzir a extrusão do veneno do narigudo e/ou veneno cuspir em alguns grupos de heteropterans. Em terceiro lugar, descrevemos os métodos para recolher as toxinas de veneno por dissecação de animais anestesiados para obter as glândulas de veneno. Este método é complementar a outros métodos, como podem permitir colheita de toxinas de táxons em que a eletroestimulação e assédio são ineficazes. Estes protocolos permitirá que pesquisadores colher as toxinas do heteropteran insetos para caracterização de estrutura-função e possíveis aplicações na medicina e na agricultura.
Introdução
Heteropteran venenos são substâncias bioativas potente1. Por exemplo, a secreção de veneno/saliva de sangue-alimentação Heteroptera como kissing bugs (Triatominae) e percevejos (Cimicidae) facilita a alimentação por perturbar a hemostasia2. Toxinas nestes venenos alvo de múltiplas vias, incluindo a coagulação, agregação plaquetária e vasoconstrição, bem como a dor e coceira caminhos. Venenos da maioria das outras espécies de heteropteran são adaptados para facilitar a predação, ao invés de sangue-alimentação. Seus venenos de causam paralisia, morte e liquefação do tecido quando injetado em invertebrados3,4. Quando injetado em animais vertebrados, seu veneno também pode ter efeitos drásticos. Por exemplo, injeção de veneno do inseto assassino Holotrichius innesi em vertebrados causa dor, paralisia muscular e hemorragia; envenomated de ratos por este bug morrer rapidamente devido a paralisia respiratória5.
Transcriptomic e proteomic estudos revelaram a composição de proteína de alguns venenos de heteropteran. Venenos de espécies de predadores são ricos em proteases, outras enzimas e peptídeos e proteínas de estrutura desconhecida e função6,7,8. Kissing bug veneno é rico em triabin proteína da família, cujos membros afetam profundamente a coagulação, agregação plaquetária e vasoconstrição2,9. No entanto, não se sabe quais toxinas subjazem a maioria dos bioactivities de veneno. Por exemplo, o veneno do kissing bug Triatoma infestans foi relatado para ser analgésico e inibe canais de sódio10, mas os componentes responsáveis continuam a ser elucidado. Da mesma forma, não se sabe qual componente ou componentes do veneno de inseto assassino causam paralisia ou dor. Um pré-requisito para identificar as responsável pela bioactivities de veneno especial e para caracterizar a estrutura e função de toxinas de veneno de romance, de toxinas é a obtenção de veneno.
Veneno foi obtido de heteropterans por eletroestimulação5,6,7,8,11,12,13, provocação de defensiva respostas4,8, mecanicamente, apertando o tórax12,14,15,16, dissecando fora as glândulas de veneno8,17 ,18,19,20,21,22e aplicação dos agonistas dos receptores de acetilcolina muscarínicos23. Julgar as potenciais vantagens e desvantagens de qualquer método é complicada pela morfologia das glândulas de veneno heteropteran, que consistem de uma glândula principal com dois lúmens separadas, a glândula principal anterior (AMG) e posterior principal glândula (PMG), bem como um associado a glândula acessória (AG). Esses compartimentos diferentes glândulas produzem secreção de proteínas diferentes, que pode ser especializada para diferentes funções biológicas, incluindo a captura de presas, defesa e digestão extra oral8,17. Em peiratine e ectrichodiine barbeiros, a AMG tem sido associada com a captura de presas e a PMG com digestão extra oral17. No entanto, no harpactorine bug Pristhesancus plagipennis o PMG é especializado para captura de presas e digestão Considerando que a AMG é a hipótese da secretar veneno defensivo8. A AG tem sido descrito como tendo pouca função secretora em barbeiros8 ou como um importante local de armazenamento de protease em bugs de água gigante23. Claramente, mais trabalho é necessário para esclarecer a função de cada compartimento de glândula entre vários subgrupos de heteropteran e para determinar a função da maioria das toxinas do veneno. Neste relatório, descrevemos protocolos para colheita de toxinas de veneno de heteropterans em direção a esse objetivo.
Protocolo
Este protocolo está em conformidade com a política da Universidade de Queensland estabelecida no cuidado responsável e uso de animais em ensino e pesquisa (PPL 4.20.11), bem como a saúde nacional e do Medical Research Council código australiano para o cuidado e uso de animais para fins científicos (8th edição 2013).
Atenção: Tome cuidado para não ser envenomated quando estiver manipulando barbeiros. Tome cuidado para proteger os olhos ao manusear espécies que cuspir veneno defensivamente. Tome cuidado ao longo para não machucar os animais experimentais. Isso inclui monitoramento de pressão às restrições tais como elásticos e garantindo que o narigudo não está quebrado.
Nota: Opcionalmente, anestesiar animais pela exposição ao CO2 para 0,5-2 min ou resfriamento de 4-10 ° C antes da colheita de veneno em visa 1-3 para facilitar a transferência segura e contenção. Anaesthetization não é estritamente necessário, mas pode facilitar a contenção segura de espécimes ágil ou fortes. No entanto, os animais devem ser acordados para permitir que o veneno da colheita. Lembre-se aplicações a jusante ao decidir se deve ou não adicionar inibidores de protease.
1. colheita de toxinas de veneno por eletroestimulação
- Obter espécimes vivos de que a colheita de toxinas.
- Pinça de plástico pré-preparadas uso com eletrodos positivos e negativos montado em qualquer dica. Ligue pinça eletrificada a uma Chatta ou uma fonte de tensão constante, o que permite o ajuste da tensão.
- Para as pequenas (~ 10 mm) e grande (~ 25mm) barbeiros, use tensões de pico de 15 e 25 V respectivamente.
- Para maiores heteropterans como bugs de água gigante, use até 40 V.
- Contenha erros ao vivo amarrando-os para uma plataforma, usando uma faixa de borracha sobre o tórax.
- Coloque a ponta do probóscide em uma dica de recolha apropriada. Para barbeiros, use uma ponta de pipeta P200. Para bugs de água gigante, corte a extremidade uma ponta de P200 para aumentar o tamanho da abertura.
- Delicadamente levante a tromba com um par fechado de uma pinça limpa e empurre a abertura da ponta da coleção sobre o fim do probóscide.
- Se desejar, água ultrapura de captação ~ 5 µ l antes de colocar a tromba para a coleta de uma dica. Isto reduz perdas de veneno, permanecendo dentro da ponta, apesar do veneno colhido será diluído.
- Aplica a eletroestimulação. Mergulhe os eléctrodos estimulantes em gel condutor, tais como 2,5 M NaCl/50% de glicerol. Aplica os eléctrodos no tórax. Para belostomatids, aplicam-se os dois eletrodos de superfície dorsal posterior da cabeça.
- Armazenar o veneno para evitar autodegradation. Depois de veneno é extrudado, transferi-lo rapidamente para um tubo a-20 ° C ou -60 ° C, ou um tubo contendo coquetel de inibidor de protease.
- Repita as etapas de 1.5 e 1.6 até veneno suficiente é adquirido ou sem veneno mais próximo.
2. a colheita de toxinas de veneno por assédio
- Preparar os animais para colheita de veneno e colocar a ponta da probóscide em um vestíbulo de coleção, conforme descrito nas subseções 1.1 e 1.3-1.4.
- Se o veneno é expulso espontaneamente, vá para o passo 2.3. Se não, assediar os animais tocando isso nas pernas, abdômen e antenas com uma pinça até veneno é produzido.
- Rapidamente transferi veneno para um tubo de-20 ° C-60 ° C, ou um tubo contendo inibidor da protease cocktail, se desejado.
3. a colheita de toxinas de veneno por assédio de "Cuspir" espécie de veneno
- Anaesthetize, ou anaesthetize em parte, o inseto antes de removê-lo do seu gabinete para evitar qualquer prematuro cuspindo na defensiva.
- Provoca o veneno cuspindo o comportamento. Conter e reposicionar o inseto usando a tampa profunda de um padrão de 90 x 16 mm placa de Petri. Segure a tampa ligeiramente posterior e 1-4 cm acima do inseto para impedir o voo. A maioria dos insetos vão cuspir várias vezes, muitas vezes em sucessão rápida. Certifique-se de que todo veneno é coletado na parte inferior do prato.
- Colete o veneno na parte inferior da caixa de Petri enxaguando com 10 µ l de água ultrapura. Rapidamente transferi-lo para um tubo a-20 ° C ou -60 ° C, ou um tubo contendo coquetel de inibidor de protease.
4. colheita de toxinas de veneno por dissecação da glândula
- Sacrificar animais. Fortemente anestesiar ou matar animais usando > 5 min exposição ao CO2. Canalize o puro CO2 diretamente para os buracos de ar do compartimento de habitação do animal.
- Inseto de PIN para bandeja de dissecação. Para barbeiros, disse através da superfície ventral (4.3). Para bugs de água gigante, disse através da superfície dorsal (4.4).
- Ventral dissecação
- Inserir três pinos do abdômen posterior para segurar o inseto sem perfurar as glândulas de veneno.
- Corte uma pequena incisão na superfície ventral do abdômen com um bisturi em miniatura. Use tesoura miniatura para estender a incisão anterior para a cabeça, tendo o cuidado de cortar o exoesqueleto apenas e não danificar estruturas internas.
- Para expor as estruturas internas, fazer vários cortes laterais, estendendo-se desde a incisão mediana para o lado do inseto. Em seguida, pino volta cada retalho de exoesqueleto ventral para revelar as estruturas internas.
- Para grandes barbeiros, fazer quatro incisões laterais, no abdômen médio abdome, anterior, entre as pernas do primeiras e segunda e na frente da perna primeira.
- Dorsal-dissecação
- Retire as asas perto da base. Inserir três pinos do abdômen posterior para segurar o inseto sem perfurar as glândulas de veneno.
- Corte uma incisão mediana da cabeça, no abdômen usando miniatura tesouras e bisturi, tendo o cuidado de cortar o exoesqueleto apenas e não danificar estruturas internas.
- Força as duas metades do inseto. Coloque vários pinos lateralmente ao longo do comprimento do inseto para deixar a cavidade interna exposta.
- Remova os músculos de voo usando uma pinça.
- Inunde a bandeja de dissecação. Adicione PBS até que o bug está submersa para permitir que estruturas internas flutuam acima e ser mais facilmente visualizado.
- Usando uma pinça e microtesoura, cuidadosamente remova conjuntivo e o tecido nervoso e traqueia. As glândulas de veneno aparecem como alongado, estruturas translúcidas, estendendo-se ao longo de cada lado do canal alimentar.
- Identifica a principal glândula pelo seu aspecto característico, com lobos anteriores e posteriores e duas condutas de reunião para o Hilo.
- Se desejado, identifica a glândula acessória ao traçar a conduta do hilo. Livre a principal glândula cortando as duas condutas que emana o Hilo.
- Colha os lúmens glândula desejado. Transferir a glândula para microcentrifuga de gelo contendo 30 µ l de PBS ou PBS e coquetel de inibidor de protease. Lance as glândulas com um pino afiada limpa.
- Vórtice de 10 s e centrífuga (1 min, 5.000 × g, 4 ° C) para esvaziar os lúmens da glândula. Remova o tecido glandular, usando uma pinça.
- Clarificar o extrato de toxina. Centrífuga (5 min, 17.000 × g, 4° C) para remover quaisquer partículas sólidas, retendo o sobrenadante e descartando a pelota. Armazenar a-20 ° C ou -60 ° C para evitar a degradação de autoproteolytic.
Resultados
Algumas espécies de heteropteran, como o harpactorine p. plagipennis e o reduviine Platymeris Radamanto, confiantemente produzem grandes quantidades (5-20 µ l) de veneno em resposta a eletroestimulação (tabela 1). Em geral, a maioria dos erros de peiratine, reduviine e harpactorine produzem veneno em resposta a esse método. Entre stenopodaine bugs, eletroestimulação suscitou veneno de Oncocephalus SP. mas não Thodelmus sp. Os erros de holoptiline e emesine amostrados não deu veneno significativo (por exemplo, suficiente para análise por espectrometria de massa) em resposta a eletroestimulação. Eletroestimulação também pode ser usada para colher a veneno de insetos belostomatid e percevejos predadores. No entanto, eletroestimulação de escorpiões de água (Nepidae) induzida liberação do conteúdo das glândulas cefálicas apenas, em vez de veneno de probóscide. Falha para colher veneno por eletroestimulação em algumas espécies é mais provável devido à complexidade morfológica de glândulas de veneno e os mecanismos fisiológicos, controlando a liberação de veneno8.
Além de liberar o veneno devido a eletroestimulação, os reduviids p. plagipennis, Havinthus rufovarius, p. Radamanto e o belostomatid distinctifemur Lethocerus, vontade espontaneamente ejetar veneno de probóscide durante a manipulação. Ejeção tal veneno é frequentemente acompanhada por exibe defensiva. P. Radamanto também cospe veneno defensivamente4, um comportamento que ocorre em cobras24 e aranhas25 mas da qual não estamos conscientes em qualquer outra espécie de reduviid.
SDS-PAGE e proteomics experimentos demonstram que os venenos colhidos por eletroestimulação e assédio são ricos em proteínas6,7,8. As proteínas são responsáveis por uma grande proporção do presente material, embora também é provável que os venenos contêm iões inorgânicos e outras substâncias. Veneno de inseto assassino obtido por eletroestimulação e assédio normalmente contém mais de uma centena de peptídeos e proteínas (Figura 1, Figura 2). Belostomatid veneno anteriormente foi relatado para ser rico em lysophospholipids13. Espectros de absorção de infravermelho de veneno de belostomatine água bug Diplonychus eques são consistentes com um teor de proteínas e lysophospholipids. Para o lethocerine L. distinctifemur, a evidência foi encontrada somente para proteínas e não lysophospholipids6.
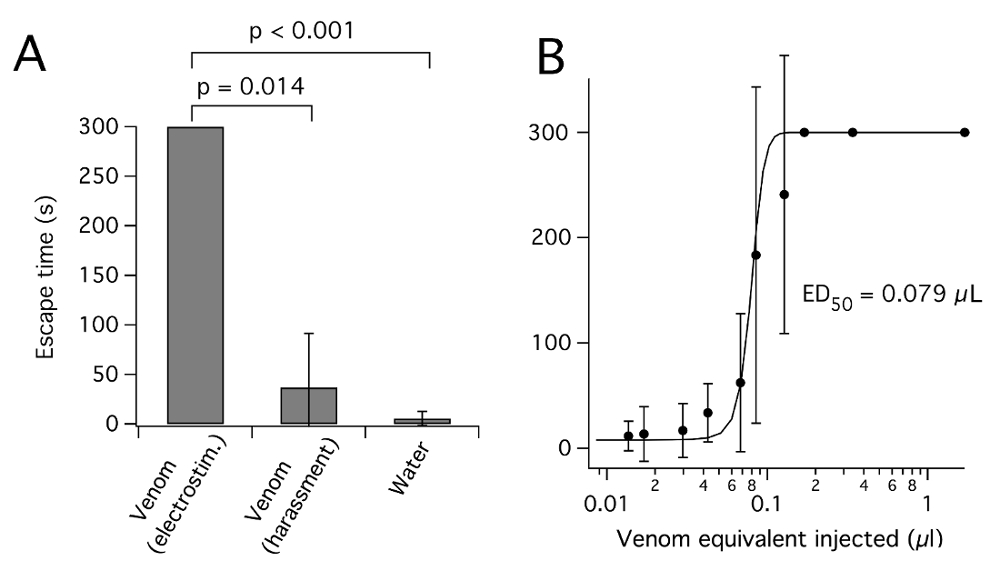
Conforme relatado por venenos de aranha26, veneno de insetos heteropteran colhido é susceptível de variar em concentração e composição, dependendo do inseto usado e o método pelo qual é colhida. Espectroscopia de UV das amostras diluídas veneno sugere valores de absorvância (280) de 50-250 (10 mm de comprimento de caminho) para veneno não diluído, consistente com uma concentração elevada de proteína de ~ 50-250 mg/mL7,12,19. Privação de rapina foi relatada para causar aumento sucessivo em veneno concentração e paralisante potencial3 , bem como reduções sucessivas no pH27. No entanto, fome prolongada resultará na perda da condição e da morte. Bem como a concentração, o método pelo qual veneno é colhido a partir heteropterans pode afetar sua composição. A composição de toxina do veneno do bug assassino p. plagipennis marcadamente diferentes dependendo se foi colhida por eletroestimulação ou assédio8. No caso de p. plagipennis, isto foi mostrado para ser devido a eletroestimulação, rendendo o conteúdo da PMG, Considerando que o assédio rendeu o conteúdo da AMG. Veneno obtido por eletroestimulação, mas não é assédio, insetos potently paralisado (Figura 3). No entanto, não está claro em que medida este resultado pode ser generalizado para outro Reduviidae ou outro Heteroptera.
Colheita de veneno diretamente por dissecando as glândulas de veneno permite que os mecanismos de controlo das glândulas de veneno a ser evitado, ao custo de contaminação com proteínas do tecido glandular (não-venom). Independentemente disso, extratos obtidos de material dissecado podem ser usados para ensaios de bioactividade/toxicidade. Por exemplo, extratos da PMG, AMG e AG de p. plagipennis, preparado usando o protocolo acima, foram analisados utilizando espectrometria de massa de cromatografia líquida/tandem8. Este processo identificou um total de 182, 114 e 71 proteínas no total, dos quais 45, 51 e 12 foram classificadas como proteínas do veneno putativo baseadas em características de sequência de aminoácidos, com as proteínas restantes classificadas como proteínas putativo das tarefas domésticas. Injeção de extratos da PMG, mas não AMG ou AG, em insetos resultou em paralisia e morte8.
| Infraordem | Família | Subfamília | Nome binomial | Nome comum | Eletroestimulação | Assédio | Dissecação |
| Cimicomorpha | Reduviidae | Harpactorinae | Pristhesancus plagipennis | Bug de assassino comum Brisbane | √ | √ | √ |
| Havinthus rufovarius | Bug do assassino de tigre vermelho | √ | √ | √ | |||
| Scipinia arenacea | Proechimys vermelho assassino bug | √ | nd | √ | |||
| Gminatus spp. | Laranja grande assassino bug | √ | nd | √ | |||
| Trachylestes aspericollis | Pequeno bug vermelho do assassino | √ | nd | nd | |||
| Reduviinae | Platymeris spp. | Inseto assassino Africano gigante | √ | √ | √ | ||
| Psytalla horrida | Proechimys bug assassino | √ | nd | √ | |||
| Peiratinae | Ectomocoris spp. | Bug de assassino de laranja da terra | √ | nd | √ | ||
| Peirates spp. | Bug de assassino preto | √ | nd | nd | |||
| Stenopodainae | Oncocephalus spp. | - | √ | nd | √ | ||
| Thodelmus spp. | - | x | nd | √ | |||
| Holoptilinae | Lêmure Ptilocnemus | Bug-de-patas-pena | x | x | nd | ||
| Emesinae | Stenolemus spp. | Bug de patas por segmento | x | x | x | ||
| Pentatomomorpha | Pentatomidae | Asopinae | Amyotea hamata | Percevejo predador amarelo | √ | nd | nd |
| Nepomorpha | Nepidae | Ranatrinae | Ranatra dispar | Escorpião da água | x, cg | x | √ |
| Belostomatidae | Belostomatinae | Diplonychus eques | Bug de água | √ | nd | nd | |
| Belostomatidae | Lethocerinae | Lethocerus sp. | Inseto gigante de água | √ | √ | √ | |
| carrapato, bem sucedido; Cruz, sem sucesso; nd, não determinado; CG, descarga de glândula cefálica apenas | |||||||
Tabela 1: Especificidade de táxon dos métodos utilizados para a colheita de veneno de heteropterans.

Figura 1 : Proteínas detectadas por LC-MS/MS análise de manchas de SDS-PAGE 2D e fracções HPLC de veneno coletados de P. plagipennis por eletroestimulação (protocolo 1), apresentando abundantes proteases, CUB-domínio proteínas e proteínas heteropteran veneno família 1. 2D gel de SDS-PAGE (A) de petróleo bruto p. plagipennis veneno, mostrando famílias da proteína identificadas por LC-MS/MS de gel de manchas. (B), HPLC cromatograma do fracionamento do veneno de p. plagipennis , mostrando famílias da proteína identificadas por LC-MS/MS análise de frações coletadas. Reproduzido com permissão7. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 : Proporção de sequências pertencentes a cada classe de proteína principal no veneno da Plagipennis p.. Reproduzido com permissão7. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 : P. plagipennis veneno obtido por eletroestimulação, mas não é assédio, paralisa insetos. (A) efeito de injetar veneno obtido por eletroestimulação ou assédio ou água, na fuga de críquete. Para cada condição de veneno, veneno de µ l 0.17 equivalente foi injetado no abdômen e o tempo para escapar de uma tampa do prato de petri arrebitado (em s, até 300 s, média ± SD) foi marcado. (B) Dose-resposta curva para inibição de sucesso escapar pelo veneno obtido a partir de p. plagipennis por eletroestimulação. Reproduzido com permissão de8. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
O passo mais crítico na colheita veneno de inseto assassino está selecionando o método apropriado dependendo os propósitos do estudo. Cada um dos três métodos apresentados para colheita heteropteran venenos tem vantagens e desvantagens dependendo de aplicações a jusante.
Indução de bugs para expelir o veneno de probóscide (protocolos de 1-3) evita a contaminação de veneno por tecidos glandulares. Além disso, esses métodos são não-letal e podem ser repetidos muitas vezes ao longo da vida de um inseto. Eletroestimulação, geralmente, fornece maiores quantidades de veneno e produz veneno com potente toxicidade a presa insetos de acordo com vários estudos5,8. Provocar uma resposta defensiva é outra maneira de extrair veneno de probóscide e um que pode render o veneno do teor de proteínas diferentes para eletroestimulação8. No entanto, eletroestimulação e provocação não funcionam para muitas espécies, e sem uma investigação paralela da saída de secreção das glândulas de veneno, não está claro qual lúmen da glândula (ou qual combinação de lumens de glândula) está sendo colhida.
Colheita de veneno por dissecção (protocolo 4) é em muitos aspectos complementares. Dissecação representa uma maneira direta de acesso veneno armazenado, e cada compartimento da glândula de veneno pode ser colhido separadamente ou em pool (ou seja, a possibilidade de que tenha sido colhido o veneno 'errado' é evitado). No entanto, o método é letal e além disso faz com que a contaminação ligeira de veneno por componentes de tecido. Heteroptera muitos são muito pequenas (ou demasiado alongada no caso Emesinae, os bichos-de-patas-segmento) para permitir que o veneno da colheita por dissecação. Se dissecação é usada para extrair proteínas dos compartimentos individuais glândula separadamente, é fundamental para separar os lóbulos rapidamente e extrair seu conteúdo separadamente, para evitar a contaminação cruzada.
Os métodos apresentados aqui precisará ser modificado dependendo da espécie particular estudou. Para coleta de veneno por eletroestimulação, os principais aspectos para otimizar são como o bug é retido. Por exemplo, a maioria dos reduviids são capazes de estender sua tromba sobre uma escala larga do movimento. Estas espécies podem ser simplesmente conteve direito-forma-acima em uma plataforma usando um elástico, e a tromba evertido manualmente. Para espécies com proboscis menos flexíveis, tais como belostomatids, em vez disso é necessário conter os insetos em uma posição de cabeça para baixo e abaixar um receptáculo de coleção no ângulo correto usando uma retorta ou braço mecânico. A magnitude e o padrão de electricidade aplicada também devem ser otimizados, e neste caso, é melhor começar baixo e lentamente aumentar a tensão aplicada para evitar a letalidade.
Se o objectivo de um estudo é alcançar uma compreensão detalhada de como uma determinada espécie produz e usa veneno, uma investigação aprofundada, combinando vários colheita métodos, bem como tecnologias como espectrometria de massa e experimentos de RNA-Seq, pode ser Necessário. Se o objectivo é usar heteropteran venenos como bibliotecas de moléculas biológicas será projectado para alguma atividade biológica desejada, em seguida, um painel de amostras de veneno obtido por eletroestimulação, assédio, e/ou dissecação pode ser adequada. No entanto, constatamos que o papel biológico normal do veneno colhido é susceptível de determinar que bioactivities estão presentes. Por exemplo, veneno usado para predação é mais provável que contêm compostos de inseticida, Considerando que o veneno usado para a defesa é mais provável que contêm agentes algogenic (causando dor).
Não incluímos a colheita de veneno por aplicação da pilocarpina de agonista do receptor de acetilcolina muscarínicos no presente protocolo. Futuros experimentos são necessários para determinar as características de expulsão de veneno induzidas pela pilocarpina em comparação com os métodos acima.
Neste artigo, apresentamos métodos que permitirão aos pesquisadores obter venenos de insetos heteropteran. Coleção de veneno bem sucedida permitirá ainda mais investigações sobre a produção, composição, função e evolução de veneno em Heteroptera. Além disso, algumas toxinas heteropteran podem encontrar o utilitário como inseticidas ecológicos, moléculas de chumbo para desenvolver terapêutica humana, ou como ferramentas científicas para investigar os sistemas biológicos.
Divulgações
Os autores não têm nada para divulgar.
Agradecimentos
Reconhecemos financeiras suporte do Conselho australiano de pesquisa (bolsas DP130103813 e LP140100832 para G.F.K., DECRA Fellowship DE160101142 para EABU), o Australian National Health & Medical Research Council (bolsa de pesquisa Principal APP1044414 para G.F.K.) e a Universidade de Queensland (Postdoctoral Fellowship para A.A.W.).
Materiais
| Name | Company | Catalog Number | Comments |
| Electostimulator | Grass Technologies | S48 Square Pulse Stimulator | Electrostimulator allowing pulsed electrostimulation |
| Featherlight tweezers | Australian Entomological Supplies | E122B | For handling live venomous insects |
| Protease inhibitor cocktail | Sigma | 4693124001 | For preventing autoproteolytic digestion of venom |
| Dissection equipment | Australian Entomological Supplies | E152Micro | For fine dissections |
| Insect pins | Australian Entomological Supplies | E162 | For fine dissections |
Referências
- Walker, A. A., Weirauch, C., Fry, B. G., King, G. F. Venoms of heteropteran insects: A treasure trove of diverse pharmacological toolkits. Toxins. 8 (2), 43 (2016).
- Ribeiro, J. M. C., Assumpção, T. C., Francischetti, I. M. B. An insight into the sialomes of bloodsucking Heteroptera. Psyche (Stuttg). 2012, 1-16 (2012).
- Ambrose, D. P., Maran, S. P. M. Quantification protein content and paralytic potential of saliva of fed and prey deprived reduviid Acanthaspis pedestris Stål (Heteroptera: Reduviidae: Reduviinae). Indian Journal of Environmental Science. 3 (1), 11-16 (1999).
- Edwards, J. S. The action and compostion of the saliva of an assassin bug Platymeris rhadamanthus Gaerst. (Hemiptera, Reduviidae). Journal of Experimental Biology. 38, 61-77 (1961).
- Zerachia, T., Bergmann, F., Shulov, A., Kaiser, E. . Animal and Plant Toxins. , 143-146 (1973).
- Walker, A. A., Hernández-Vargas, M. J., Corzo, G., Fry, B. G., King, G. F. Giant fish-killing water bug reveals ancient and dynamic venom evolution in Heteroptera. Cellular and Molecular Life Sciences. , (2018).
- Walker, A. A., et al. Giant fish-killing water bug reveals ancient and dynamic venom evolution in Heteroptera. Cell. Mol. Life Sci. , (2018).
- Walker, A. A., et al. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nature Communications. 9 (1), 755 (2018).
- Hernández-Vargas, M. J., Santibáñez-López, C. E., Corzo, G. An insight into the triabin protein family of American hematophagous reduviids: Functional, structural and phylogenetic analysis. Toxins. 8 (2), 44 (2016).
- Dan, A., Pereira, M. H., Pesquero, J. L., Diotaiuti, L., Beirao, P. S. Action of the saliva of Triatoma infestans (Heteroptera: Reduviidae) on sodium channels. Journal of Medical Entomology. 36 (6), 875-879 (1999).
- Corzo, G., Adachi-Akahane, S., Nagao, T., Kusui, Y., Nakajima, T. Novel peptides from assassin bugs (Hemiptera: Reduviidae): isolation, chemical and biological characterization. FEBS Letters. 499 (3), 256-261 (2001).
- Sahayaraj, K., Kumar, S. M., Anandh, G. P. Evaluation of milking and electric shocks for venom collection from hunter reduviids. Entomon. 31 (1), 65-68 (2006).
- Silva-Cardoso, L., et al. Paralytic activity of lysophosphatidylcholine from saliva of the waterbug Belostoma anurum. Journal of Experimental Biology. 213 (19), 3305-3310 (2010).
- Noeske-Jungblut, C., et al. Triabin, a highly potent exosite inhibitor of Thrombin. Journal of Biological Chemistry. 270 (48), 28629-28634 (1995).
- Noeske-Jungblut, C., et al. An inhibitor of collagen-induced platelet aggregation from the saliva of Triatoma pallidipennis. Journal of Biological Chemistry. 269 (7), 5050-5053 (1994).
- Sahayaraj, K., Borgio, J. F., Muthukumar, S., Anandh, G. P. Antibacterial activity of Rhynocoris marginatus (Fab.) and Catamirus brevipennis (Servile) (Hemiptera: Reduviidae) venoms against human pathogens. Journal of Venomous Animals and Toxins Including Tropical Diseases. 12 (3), 487-496 (2006).
- Haridass, E. T., Ananthakrishnan, T. N. Functional morphology of the salivary system in some reduviids (Insecta-Heteroptera-Reduviidae). Proceedings of the Indian Academy of Sciences. Animal Sciences. 90 (2), 145-160 (1981).
- Maran, S. P. M., Ambrose, D. P., Ignacimuth, A., Sen, A., Janarthanan, S. . Biotechnological Applications for Integrated Pest Management. , 125-131 (2000).
- Maran, S. P. M., Selvamuthu, K., Rajan, K., Kiruba, D. A., Ambrose, D. P., Ambrose, D. P. . Insect Pest Management, A Current Scenario. , 346-361 (2011).
- Pereira, M. H., et al. Anticoagulant activity of Triatoma infestans and Panstrongylus megistus saliva (Hemiptera/Triatominae). Acta Tropica. 61, 255-261 (1996).
- Ribeiro, J. M., Marinotti, O., Gonzales, R. A salivary vasodilator in the blood-sucking bug, Rhodnius prolixus. British Journal of Pharmacology. 101 (4), 932-936 (1990).
- Ribeiro, J. M., Schneider, M., Guimarães, J. A. Purification and characterization of prolixin-S (nitrophorin 2), the salivary anticoagulant of the blood-sucking bug Rhodnius prolixus. Biochem Journal. 308 (1), 243-249 (1995).
- Swart, C. C., Deaton, L. E., Felgenhauer, B. E. The salivary gland and salivary enzymes of the giant waterbugs (Heteroptera; Belostomatidae). Comparative Biochemistry and Physiology A Molecular & Integrative Physiology. 145 (1), 114-122 (2006).
- Rasmussen, S., Young, B., Krimm, H. On the 'spitting' behaviour in cobras (Serpentes: Elapidae). Journal of Zoology. 237 (1), 27-35 (1995).
- Fink, L. S. Venom spitting by the green lynx spider, Peucetia viridans (Araneae, Oxyopidae). Journal of Arachnology. 12, 372-373 (1984).
- Herzig, V. Ontogenesis, gender, molting influence the venom yield in the spider Coremiocnemis tropix (Araneae, Theraphosidae). Journal of Venomous Research. 1, 76-83 (2010).
- Sahayaraj, K., Subramanium, M., Rivers, D. Biochemical and electrophoretic analyses of saliva from the predatory reduviid species Rhynocoris marginatus (Fab). Acta Biochimica Polonica. 60 (1), 91-97 (2013).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados