
Method Article
Particulas de Lentivirus mediada produção de camundongos transgênicos: um método simples e altamente eficiente para estudo direto dos fundadores
Neste Artigo
Resumo
Aqui, apresentamos um protocolo para promover a integração do transgene e produção de ratos transgénicos fundador com eficácia elevada por uma simples injeção de um vetor de Lentivirus no espaço de um oócito fertilizado perivitelline.
Resumo
Há quase 40 anos, injeção de DNA pró-nuclear representa o método padrão para gerar ratos transgénicos com integração aleatória de transgenes. Um procedimento de rotina é amplamente utilizado em todo o mundo e sua principal limitação reside na eficácia da integração do transgene, resultando em um baixo rendimento dos animais fundador pobre. Apenas alguns por cento dos animais nascidos após a implantação de oócitos fertilizados injetados integraram o transgene. Em contraste, Lentivirus vetores são ferramentas poderosas para a transferência de genes integrativa e sua utilização para transduce oócitos fertilizados permite uma produção altamente eficiente do fundador ratos transgénicos com um rendimento médio acima de 70%. Além disso, toda a tensão do mouse pode ser usada para produzir animais transgénicos e a penetrância da expressão do transgene é extremamente elevada, acima de 80% com particulas de Lentivirus transgênese mediada comparado a microinjeção de DNA. O tamanho do fragmento de DNA que pode ser a carga pelo vetor de Lentivirus é limitado a 10 kb e representa a principal limitação deste método. Usando um simples e fácil de executar o procedimento de injeção por baixo da zona pelúcida de oócitos fertilizados, mais de 50 animais do fundador podem ser produzidos em uma única sessão da microinjeção. Tal método é altamente adaptado para executar, diretamente em animais fundador, ganho rápido e perda de estudos de função ou de regiões de DNA genômicas tela por sua capacidade de controlar e regular o gene expressão na vivo.
Introdução
O trabalho pioneiro de Gordon et al . em 1980 mostrou que, após a implantação em ratos pseudopregnant, a injeção de DNA de plasmídeo para o sexo masculino pró-núcleos de oócitos fertilizados pode render a produção de animais transgénicos que integrado a Plasmideo DNA1. A demonstração de que os mamíferos transgênicos podem ser gerados teve um enorme impacto sobre ciências da vida globais, abrindo o caminho para novos campos de pesquisa, tanto para as ciências básicas e ciências biomédicas translacionais. Nas últimas quatro décadas, a microinjeção de DNA tornou-se uma prática de rotina. Embora um enorme número de ratos transgênicos foram produzido, o método padrão não é totalmente utilizável para todas as estirpes de rato e requer demorado retrocruzamentos2,3. Sua aplicação a outras espécies permanece desafiador4 e o rendimento global de integração do transgene é limitado a alguns porcentagem de animais nascido5. Além disso, a eficácia da integração do transgene representa o fator limitante que explica o pobre rendimento total da injeção de DNA pró-nuclear. A este respeito, Integrativa vetores virais são as ferramentas mais eficientes para carga e integram transgenes em assim poderiam fornecer novos meios para aumentar significativamente o rendimento de integração, a única limitação é que o tamanho do transgene que não pode exceder 10 kb6 .
Particulas de Lentivirus vetores pseudo digitados com a proteína do envelope do vírus da estomatite Vesicular (VSV) são ferramentas de transferência do gene pantropic e altamente integrativa e podem ser usados para transduce oócitos fertilizados7. A zona pelúcida envolvendo oócitos é uma barreira natural de vírus e precisa ser passado para permitir a transdução com os vetores de Lentivirus. Animais transgénicos foram gerados pelo transducing oócitos fertilizados após micro perfuração ou remoção da zona pelúcida8,9. No entanto, injeção sob a zona pelúcida no espaço perivitelline parece ser o método mais simples para transduce os ovos fertilizados como inicialmente descrito por Lois e colegas7.
A injeção de perivitelline de Lentivirus vetores permite altos rendimentos na produção de animais transgénicos que estão acima de 70% dos animais nascidos. Tal rendimento é mais 10-fold maior do que o melhor rendimento que pode ser conseguido usando padrão pró-núcleos DNA injeção7,10,11. Neste contexto, uma única sessão de injeções irá gerar pelo menos 50 fundadores transgénicos (F0). O grande número de fundadores é, portanto, compatível com fenotipagem do transgene efeito diretamente realizada em F0 ratos sem a necessidade de gerar linhas de rato transgénico. Esta vantagem permite a seleção rápida do efeito do transgene e é especificamente adaptada para executar na vivo ganho e perda de estudos de função dentro de semanas. Além disso, elementos reguladores do DNA podem também ser rapidamente analisados para mapear enhancers e motivos de DNA vinculados por fatores de transcrição11,12. Com injeções pró-nuclear, transgenes integrar normalmente como várias cópias em um único locus. Com vetores de Lentivirus, integração ocorre em múltiplos loci como uma única cópia por locus10,13. Portanto, a multiplicidade dos loci integrado é provavelmente associada a penetrância de expressão muito alta observada em fundadores transgénicos, que faz com que o novo modelo gerado mais robusto.
Importante, quando usando injeção pró-nuclear do ADN, visualização dos pró-núcleos durante o procedimento é absolutamente necessária. Essa limitação técnica impede o uso de oócitos fertilizados, provenientes de uma grande variedade de estirpes de rato. Portanto, a produção de um modelo transgênico em uma determinada tensão para qual pró-núcleos são invisíveis exige a produção de animais em uma cepa permissiva, seguido pelo menos 10 sucessivos retrocruzamentos para transferir o transgene no mouse desejado estirpe. Com as injeções de vetor de Lentivirus, perivitelline espaço é sempre visível e a injeção não requer habilidades altamente específicas. Por exemplo, ratos transgénicos de NOD/SCID que não são apropriados para injeção de pró-núcleos foram obtidos com o vector viral injeções14.
Aqui, apresenta-se um protocolo abrangente para permitir que a simples produção de camundongos transgênicos usando injeções de vetor de Lentivirus no espaço perivitelline de um embrião de fase de uma célula. Expressão do transgene controlada com qualquer onipresente ou promotores específicos de célula é descrito em detalhes.
O backbone de Lentivirus de ΔU3 de pTrip foi usado no presente estudo15. Esse vetor permite produzir vetores de Lentivirus defeituoso replicação, no qual a sequência de U3 foi parcialmente excluída para remover a atividade de promotor U3 e gerar um self inactivating vector (SIN)16. Estoques de vetor de Lentivirus foram produzidos por transfecção transiente de células HEK-293T com o p8.91 encapsulamento do plasmídeo (ΔVpr ΔVif ΔVpu ΔNef)6, a codificação a estomatite vesicular (VSV) de vírus glicoproteína-G17e o ΔU3 de pTRIP pHCMV-G vector recombinante. O processo de produção detalhado é fornecido como métodos complementares.
Produção das unidades populacionais de vetor de Lentivirus de alta concentração é realizada em condições de biossegurança nível II (BSL-2). Isto é verdade para a maioria dos transgenes exceto oncogenes que têm de ser produzidos em BSL-3. Portanto, a produção em condições de BSL-2 para a maioria dos casos é suficiente. Além disso, o uso e a produção são geralmente desconectados para a maioria das agências reguladoras nacionais, lidando com organismos geneticamente modificados (OGM). Quantidades limitadas de replicação incompetente SIN Lentivirus vetores (abaixo de 2 µ g da proteína do capsídeo p24) podem ser usadas sob condições de BSL-1 conforme descrito pela agência francesa OGM, de acordo com as recomendações da União Europeia.
Protocolo
Todos os procedimentos que incluem trabalho animal têm aprovação ética e autorizados pelo Ministério francês de investigação e educação sob o número APAFIS #5094-20 16032916219274 v6 e 05311.02. As instalações de animais ICM PHENOPARC foi credenciado pelo Ministério da agricultura francês sob o número de acreditação B75 13 19. O protocolo global necessita de executar cada procedimento dentro de um frame de tempo preciso que é resumido na Figura 1.
1. animal compra e preparação de compostos básicos
- Compra de animais
- Ordem de 25 homens vasectomizados B6CBAF1/JRj que são 8 semanas de idade (geração F1 original cruza entre ♀C57Bl/6JRj e ♂CBA/JRj).
Nota: Isolar os machos à chegada. Vasectomizado machos podem ser reutilizados pelo menos um ano.
Mude as gaiolas em 3 semanas. - Encomendar 50 B6CBAF1/JRj fêmeas que são 10 semanas de idade e mantém uma piscina de pelo menos 50 animais.
- Ordem de 10-15 C57BL/6JRj machos férteis que são 8 semanas de idade.
- Ordem 30 C57BL/6JRj as fêmeas que são 4 semanas de idade.
- Ordem de 25 homens vasectomizados B6CBAF1/JRj que são 8 semanas de idade (geração F1 original cruza entre ♀C57Bl/6JRj e ♂CBA/JRj).
- Anestesia e eutanásia
- Anestesia é realizada utilizando um volume de mistura de xilazina/cetamina 300μL, injectado intraperitonealmente (cetamina na dose de 150μg/g de massa corporal, xilazina, na dose de 0.15μg / g de peso corporal). Os animais são colocados sob aquecimento pad para ajustar a temperatura do corpo. Verificar os reflexos por beliscar a cauda do animal antes de iniciar o procedimento.
- A eutanásia foi realizada por deslocamento cervical. Decapitação foi incluída como um método secundário para confirmar a morte do animal. Eutanásia de embriões foi realizada por decapitação.
Nota: Após a chegada, permitem que os animais um mínimo de 1 semana para se habituar a instalação (sem manipulação ou acasalamento). Importante, qualquer cepas de rato incluindo linhas transgénicas podem ser utilizadas como férteis machos e fêmeas férteis para superovulação. A escolha da variedade deve ser feita de acordo com os requisitos da questão científica.
- Preparação de hormônio
- Adicionar 910 µ l de tampão PSMG (soro de égua grávida gonadotropina) em 1 frasco de liofilizado PMSG, fazer 100 alíquotas µ l e armazenar a-20 ° C.
Nota: Cada alíquota contém 55 UI para 11 ratos. Nunca mantenha PMSG alíquotas para mais de 2 semanas após a primeira utilização. - Adicione-se frasco hCG liofilizado, 2730 µ l de tampão de hCG (gonadotrofina coriônica humana) em 1. Faça 100 alíquotas µ l e armazenar a-20 ° C.
Nota: Cada alíquota contém 55 UI para 11 ratos.
- Adicionar 910 µ l de tampão PSMG (soro de égua grávida gonadotropina) em 1 frasco de liofilizado PMSG, fazer 100 alíquotas µ l e armazenar a-20 ° C.
- Preparação de hialuronidase
- 1 frasco de hialuronidase com 3 mL de meio de M2 para obter um 10 mg/mL de solução-mãe e fazer 50 alíquotas µ l de reconstituir. Em seguida, armazenar a-20 ° C.
- Preparação de instrumentos de cirurgia
- Esterilize todos os equipamentos de cirurgia, usando o autoclave.
2. superovulação de doadoras
- Em uma instalação animal usando 12 h dia - ciclos de noite, injetar PMSG às 14:00 no dia -3. Injete hCG às 12 da manhã de dia -1 e companheiro com os machos férteis logo após injeção de hCG.
- Dia -3, adicionar 1 mL de solução de NaCl 0,9% estéril em 1 alíquota de 100 µ l de PMSG (55 UI). Injecte 10 fêmeas C57BL/6JRj com 100 µ l intraperitonealmente, usando uma seringa sem qualquer volume morto.
Nota: Cada rato receberá 5 UI de PMSG. - Dia -1, adicione 1 mL de solução de NaCl 0,9% estéril em 1 alíquota de 100 µ l de hCG (55 UI). Injete os ratos que receberam a injeção de PMSG com 100 µ l de solução diluída de hCG (5UI) intraperitonealmente. Use uma seringa sem qualquer volume morto. Executar a injeção lentamente e esperar antes de retirar a agulha, para que o líquido não vaza.
Nota: Cada rato receberá 5 UI de hCG. Injeção de hCG deve ser realizada 46 h após PMSG. - Coloque cada fêmea C57BL/6JRj na gaiola do macho garanhão diretamente após a injeção de hCG.
- Verifique plugues vaginais na manhã do dia 0 e usar as fêmeas positivas para coletar ovos fertilizados.
3. preparar as fêmeas Pseudopregnant B6CBAF1/jRj
- Acasalar com um macho vasectomizado o dia antes da coleta do ovo (dia -1) com 2 fêmeas de B6CBAF1/JRj às 17:00.
Nota: É muito importante se acasalar com as fêmeas que são provenientes de gaiolas diferentes para evitar a sincronização de ciclos femininos. Isto irá aumentar o rendimento da obtenção de tampões vaginais. Além disso, não adicione uma fêmea para uma gaiola masculina que foi alterada nos últimos 2 dias. Eficácia do comportamento reprodutivo no sexo masculino é maior quando a gaiola é suja.
4. fertilizado ovos coleção
- Preparação
- Adicione µ l 1450 de M2 a solução para preparar a solução de trabalho de hialuronidase hialuronidase.
- Coloque uma gota de 100 µ l de solução de trabalho de hialuronidase por fêmea, usada para produzir ovos fertilizados em uma placa de Petri de 100mm e manter à temperatura ambiente.
- Adicione 500 µ l do M16 em placas de 4 poços. Use 2 poços por tipo de Lentivirus vetores que será injetado: um bem irá conter os ovos injetados e o outro os que não injectada. Coloque as 4 placas bem na incubadora a 37 ° C com 5% CO2 atmosfera.
- Prepare-se pipetas para coleta e manipulação de embriões.
- Suavizar os capilares de hematócrito de vidro (75 mm/60 µ l), rodando o centro do tubo capilar de vidro duro na chama.
- Remova os capilares do calor tão rapidamente quanto possível e puxe-o para obter um tubo com diâmetro interno de cerca de 300 µm. Puxe o tubo de refrigeração para obter um tempo puro.
- Recolha os ovidutos.
- Eutanásia em fêmeas C57BL/6JRj por deslocamento cervical às 09:00 no dia 0. Morte confirmada por decapitação.
Nota: Este método de eutanásia foi aprovado pelo IACUC e segue as recomendações europeias. - Realize uma grande incisão horizontal para abrir a cavidade abdominal com uma tesoura. O oviduto situa-se entre o útero e o ovário.
- Remova o mesometrium e a membrana carregando proeminentes vasos sanguíneos com pinça curvada.
- Separe o oviduto do ovário com pinça curvada.
- Use pinça curvada como um guia para cortar o oviduto do ovário usando tesouras curvas.
- Puxar o oviduto e corte do útero com uma tesoura curva.
Cuidado: Não toque na ampola inchada que contém os ovos fertilizados. Execute todo o procedimento usando instrumentos esterilizados. - Coloque todos recolhidos ovidutos M2 médio (prato de cultura de 35 mm) à temperatura ambiente
- Coloque 2 ovidutos na mesma gota de 100 µ l de solução de trabalho de hialuronidase (0,3 mg/mL).
- Eutanásia em fêmeas C57BL/6JRj por deslocamento cervical às 09:00 no dia 0. Morte confirmada por decapitação.
- Remova células cumulus de ovos fertilizados.
- Sob um estereomicroscópio, utilize 2 seringas de insulina: o primeiro que segure o oviduto e o segundo para rasgar a ampola e dispersar fertilizado ovos para a solução de trabalho de hialuronidase.
- Levar a pipeta de vidro preparado para a coleta de ovos e ligue-o para o tubo e um filtro de 0,22 μm montado no bocal para aspirar todos os ovos. Recolher todos os ovos e lavá-los pela passagem sucessiva em 6 diferentes gotas de 100 µ l de meio M2.
- Coloque os ovos fertilizados lavados na incubadora umidificado 37 ° C, com uma atmosfera de 5% de CO2 em meio de M16.
5. fazer injeção pipetas
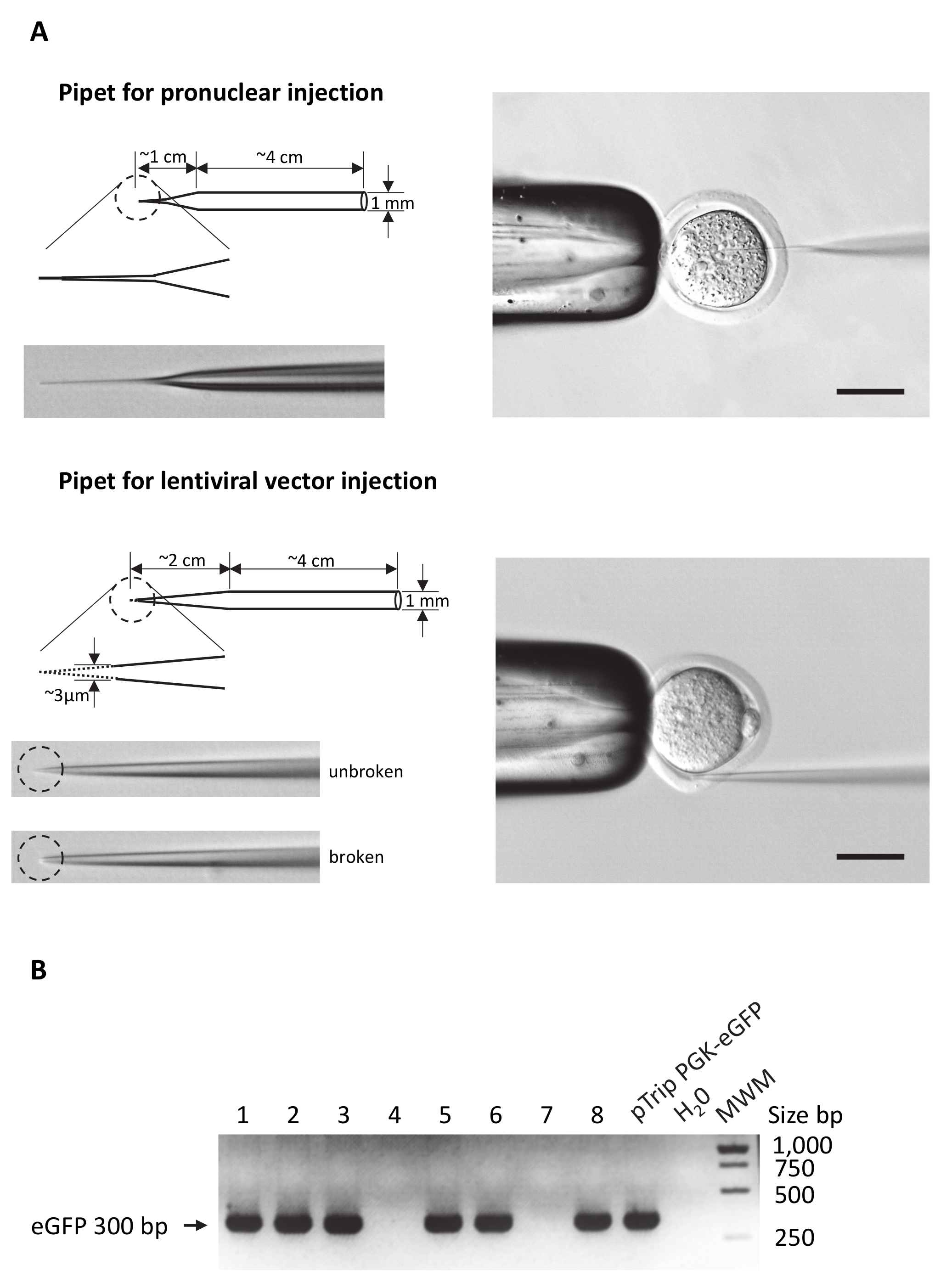
- Use o tubo capilar de vidro de paredes finas (10-15 cm de comprimento) com um diâmetro externo de 1 mm e fixar este capilar no extrator micropipeta horizontal.
Nota: Em extratores de pipeta mais horizontais, 3 parâmetros podem ser ajustados: calor energia, força e tempo de atraso entre aquecimento e puxando a puxar. Ajuste esses parâmetros para obter injetando pipetas que se assemelha o um apresentado na Figura 2A. Para os usuários que são rotineiramente realizando microinjeção de DNA, use as configurações padrão e ajustar o atraso entre aquecimento e puxando para alterar a forma global da ponta da pipeta.
6. fazer exploração pipetas
- Use uma pipeta injetando para preparar a pipeta de exploração.
- Anexe uma pipeta injetando um microforge. Corte a pipeta com o microforge para obter uma ponta aguda simétrica de 80 a 100 µm de diâmetro. Então polir a ponta com calor sobre o microforge para obter uma forma redonda simétrica sem sebes afiadas.
7. preparação da pipeta de injeção que contém o vetor de Lentivirus
- Centrifugar a suspensão de vetor de Lentivirus 160 x g por 2 min para detritos frequentemente presentes nas existências de Lentivirus congeladas de Pelotas.
- Recuperar o sobrenadante e transferir para um novo tubo de 0,5 mL em uma classe de segurança II.
- Transferi 1 µ l do sobrenadante para uma pipeta de injeção, preparada conforme descrito na etapa 5, usando um Microcarregador.
- Defina a pipeta de injeção sobre o titular do instrumento de micromanipulador o certo. Conectar-se a pipeta de exploração para a esquerda micromanipulador.
Nota: O título de transdução do vetor de Lentivirus vai ser diretamente correlacionado com a eficácia da produção do fundador. De alta eficácia (> 70%), uso de vetores virais com um título na faixa de 100 ng de p24 capsídeo proteínas / µ l. O título é expressa em unidades de transdução (TU), o título deve ser acima de 109 TU/mL. Estoques de vetor de Lentivirus devem ser produzidos por transfection transiente de 293T células com o plasmídeo de encapsidation p8.91, pHCMV-G, codificação da estomatite vesicular (VSV) de vírus glicoproteína-G, conforme descrito em métodos complementares18.
8. microinjecção
- Dispense 8 µ l de meio M2 no centro de uma depressão slide e tampa com óleo parafínico leve (embriões testados) para evitar a evaporação.
- Lugar 20 ovos à gota como menos dispersos quanto possível.
Cuidado: Não faça bolhas quando depositar os embriões. - Certifique-se de que a ponta da pipeta de injeção é aberta. Se não, toque a pipeta de injeção com a pipeta de exploração.
- Definir o microinjector para um tempo de injeção de 20 s.
Nota: A viscosidade da suspensão viral permite visualização clara da dispersão do vetor viral no espaço perivitelline. A pressão de injeção deve ser ajustada a fim de preencher todo o espaço dentro de 20 s de injeção, que representa um volume de 10 a 100 pl. Pressão de injeção não deve exceder 600 hPa. - Aspire um óvulo fertilizado que contém 2 núcleos pro e 2 corpos polares com a pipeta de exploração sob o microscópio.
- Injete o ovo com o microinjector usando as configurações descritas em 8.4, no espaço perivitelline.
Cuidado: Não toque a membrana plasmática com pipeta de injeção. - Injetar tudo fertilizados ovos disponíveis em lotes de 20 ovos e colocar os ovos injetados imediatamente médio pré-aquecido e M16 da incubadora umidificado 37 ° C, com uma atmosfera de 5% de CO2.
Nota: Incube os ovos injetados por um período mínimo de 30 min após a injeção antes de transferências de embrião.
9. transferência de embriões em fêmeas Pseudopregnant B6CBAF1/JRj
- Verificar plug cópula 16 h após o acasalamento as fêmeas B6CBAF1/JRj com machos de vasectomia B6CBAF1/JRj. Faça isso antes de iniciar a coleta de ovos.
- Prepare-se pipetas de implantação do embrião injetado.
- Fazer pipetas de implantação de embriões conforme descrito para coleta e manipulação de embriões (etapa 4.2). Selecione pipetas com diâmetro interno de cerca de 150 µm com uma parte estreita em torno de 4-5 cm de comprimento.
Nota: A dica deve ser chama lustrada, a fim de reduzir possíveis danos para os ovos ou o oviduto.- Encha com óleo parafínico leve (embrião testado) logo acima do ombro da pipeta.
- Aspire a uma bolha de ar pequena, então M2 médio e, finalmente, uma segunda bolha de ar.
- Elaborar os embriões um atrás de outro para minimizar o volume total do meio que será injetado no oviduto juntamente com os embriões.
- Acaba por carregar uma gota muito pequena de óleo parafínico leve (embriões testados) de largura de cerca de um embrião.
Atenção: Cuidado durante o manuseio da pipeta.
- Fazer pipetas de implantação de embriões conforme descrito para coleta e manipulação de embriões (etapa 4.2). Selecione pipetas com diâmetro interno de cerca de 150 µm com uma parte estreita em torno de 4-5 cm de comprimento.
- Transferência de embriões.
- Esterilize todos os instrumentos.
- Anestesia a fêmea usando uma injeção intraperitoneal de 300 μL de solução estéril de NaCl 0,9% contendo 150 μg por g de peso corporal de cetamina e 0,15 μg por g de peso corporal de xilazina.
- Verificar a profundidade da anestesia por beliscar a cauda do animal com fórceps e injetar subcutaneouly 0,1 mg/kg de analgésico (buprenorfina) antes de iniciar o procedimento.
- Barba de 2 cm em ambos os lados da parte traseira ao longo da medula espinhal ao nível da última costela.
- Coloque o mouse feminino sobre uma almofada de aquecimento e em um campo estéril. Corte uma janela de 2 x 2 cm no meio da parte traseira do mouse.
- Aplicar uma solução anti-séptica (10% iodeto de povidona) sobre a pele e fazer uma incisão transversal de 1 cm com uma tesoura e, em seguida, deslize a pele lateralmente até o ovário (cor laranja) é visível através da parede do corpo.
- Faça uma incisão de 5 mm na parede do corpo, logo acima do ovário com a tesoura bem sob um microscópio binocular cirúrgico.
- Buscar a almofada de gordura com uma braçadeira de buldogue atraumática e puxe para fora do ovário, oviduto e parte superior do útero.
- Visualize a ampola e fazer um hemisection com tesoura vannas no segmento oviduto que liga o ovário para a ampola.
- Introduzir a pipeta de transferência embrião e entregar os ovos para a ampola, parando na primeira bolha de ar na pipeta a implantação.
- Repita o procedimento na segundo oviduto.
- Perto da pele com clipes de ferida.
- Lugar do animal na recuperação de câmara (39 ° C, 30-60 min) até totalmente acordado.
- Repita a injeção analgésica após 12 h e 48 h no caso de sinais de dor ou sofrimento.
- Remova os clipes de ferida 7-10 dias após a cirurgia.
- Fêmeas de seleção implantada para a gravidez, seguindo a curva de peso em 3 dias após o implante. Um ganho de peso significativo pode ser observado de 10 a 12 dias após o implante e será indicativo para a gravidez.
Nota: Todos os embriões que desenvolverão aqui vai representam putativos fundadores transgénicos. O fenótipo destes fundadores pode ser analisado em quaisquer fases do desenvolvimento, ou após o nascimento, de acordo com a questão científica ligados à geração destes animais transgénicos.
10. genotipagem fundadores transgénicos
- Preparar a solução tampão de genotipagem com 10 mM Tris-HCl, pH 8; 5 mM EDTA, pH 8.0 com SDS de 0,2% (p/v), 50 mM de NaCl. Esterilizar o buffer de genotipagem através de um filtro de 0,22 µm e armazenar em temperatura ambiente por vários meses.
- Colocar as membranas extraembryonic (para embriões) ou pequeno pedaço da cauda (para animais nascidos) em 500 µ l do filtrado buffer de genotipagem e adicionar 15 µ l de proteinase K (20 mg/mL). Incubar durante uma noite a 55 ° C.
- Centrifugue o lisado a 15.000 x g por 5 min e em seguida, use 1 µ l do sobrenadante para a reação de PCR. Lisado pode ser armazenado a 4 ° C por vários meses.
- Execute a amplificação por PCR de um fragmento do transgene em um volume de reação 20 µ l contendo 1 tampão de x PCR, 1,5 mM MgCl2, 200 µM de dNTPs, 0,2 µM cada primers de PCR, 1 UI de Taq DNA polimerase e 1 µ l de cada amostra digerida. Como um controle negativo, use 1 µ l de H2O. Como um controle positivo, use o DNA do plasmídeo vetor de Lentivirus contendo o transgene.
- Para a deteção de eGFP descreveu o uso:
eGFP em frente da primeira demão: 5' GACCACATGAAGCAGCACGACTTCT 3'
eGFP Primer Reverse: 5' TTCTGCTGGTAGTGGTCGGCGAGCT 3' - Executar a amplificação por PCR em um thermocycler: 4 min a 94 ° C, seguido de 35 ciclos de 1 min a 94 ° C, a 1 min a 60 ° C e 2 min a 72 ° C.
- Carrega o produto do PCR em um gel de agarose 2% a fim de Visualizar um 300 bp eGFP produto PCR conforme ilustrado na Figura 2B.
Nota: Todos os indivíduos apresentando uma banda PCR bp 300 integraram o transgene eGFP e podem ser considerados como transgênicos. - Analise a expressão do transgene em animais transgénicos. Por exemplo, executar histológica e imunocoloração como ilustrado na Figura 3 e Figura 4 e descrito em métodos complementares.
Nota: Ambos análise de fenótipo e análise da expressão do transgene devem ser realizadas utilizando métodos pertinentes de acordo com a questão científica global.
11. a quantificação do número de cópia do Transgene
- Prepare as amostras de DNA por PCR quantitativo.
- Extrair DNA genômico (gDNA) da Proteinase K lisado obtido na etapa 10.3 usando um kit comercial de acordo com as instruções do fabricante.
- Quantificar gDNA por espectrofotometria em 260 nm.
- Dilua cada amostra gDNA a uma concentração final de 10 ng / µ l.
- Para cada amostra, preparar diluições em série 5 (1:5) em H2O para obter 6 tubos nas seguintes concentrações: 10 ng / µ l, 2 ng / µ l, 0,4 ng / µ l, 0,08 ng / µ l, 0,016 ng / µ l e 0.0032 ng / µ l
- Prepare a mistura de reação de PCR (qPCR) quantitativa.
- Prepare a mistura de cartilha para qPCR. Para cada par de primer usar para qPCR, adicionar 10 µ l de primer para a frente (solução de primer 100 µM), 10 µ l de primer reverso (100 µM) e 80 µ l de H2O.
Nota: Para amplificar eGFP, use primers para a frente TCCAGGAGCGCACCATCTTCTTCA e TTGATGCCGTTCTTCTGCTTGTCG de reverter. Gene Cdx2 é usado como normalizador para o qPCR (2 cópias por genoma). Para Cdx2 normalizador usar primers para a frente GCCAGGGACTATTCAAACTACAGG e inverter GACTTCGGTCAGTCCAGCTATCTT - Prepare 2 misturas de qPCR, uma com a mistura de cartilha de eGFP e um com mistura de cartilha Cdx2. Prepare a mistura de qPCR suficiente para amplificar as 6 diluições em duplicatas de cada gDNA. Uma reação de qPCR contém 3.8 µ l de H2O, 5 µ l de fluorescente verde 2 x mistura de reação e 0,2 µ l de mistura da primeira demão.
Nota: Para cada animal transgênico testar, 24 qPCR reações serão executadas. A mistura de reação é fornecida por uma máquina de 384 qPCR bem. - Para cada animal testar, distribuir: 12 poços com 9 µ l de mistura de qPCR eGFP e 12 poços com 9 µ l do mix de qPCR Cdx2. Adicione 1 µ l de cada diluição gDNA 2 poços contendo a mistura de qPCR eGFP e 2 poços contendo a mistura de qPCR eGFP.
- Deixe 2 poços para cada qPCR misturam no qual gDNA foi substituir por H2O como controlo negativo.
- Prepare a mistura de cartilha para qPCR. Para cada par de primer usar para qPCR, adicionar 10 µ l de primer para a frente (solução de primer 100 µM), 10 µ l de primer reverso (100 µM) e 80 µ l de H2O.
- Coloque a placa bem 384 na máquina de qPCR e aplicar o seguinte protocolo de execução: 10 min a 95 ° C depois de 50 ciclos de 10 s a 95 ° C e 1 min a 60 ° C.
- Analise os dados. Para cada gDNA testar, plotar os valores de Ct em função do Log do montante total gDNA (6 pontos em duplicatas). Ajustar a curva usando a regressão linear com o método menos quadrado e extrapolar o valor de Ct correspondente a interseção com o eixo y. Use os valores de Ct extrapolados para eGFP e o Cdx2 normalizado para calcular o número de cópia eGFP relativo Cdx2 (2 cópias) usando o método de padrão 2ΔdCt 11.
Resultados
Animais transgénicos foram gerados usando o protocolo aqui apresentado. Representante resultados ambos onipresente e expressão de transgene específica de tipo de célula são ilustrados.
Expressão constitutiva de transgenes
Promotores onipresentes são ferramentas de pesquisa básica para expressar transgenes de forma sustentada e eficiente. Esses promotores são usados para uma variedade muito grande de aplicações desde em vitro transfection da pilha de transgênese em vivo em pequenos e grandes animais.
Vetores de Lentivirus foram construídos para expressar o gene verde fluorescente repórter (eGFP) sob o controle do promotor o citomegalovírus (CMV) ou o promotor composto que CAG com base na fusão de promotor de actina a galinha e o realçador de CMV. Ambos os vetores Lentivirus foram produzidos (métodos complementares) e a titulação foi determinado em 293T células como unidades de transdução (TU) baseadas na expressão de eGFP. Ambas as construções do vetor de Lentivirus foram injetadas no espaço perivitelline de oócitos fertilizados em uma concentração de 109 TU/mL e implantado em ratos fêmeas grávidos pseudo. Embriões implantados próximo foram coletados apenas antes do nascimento e genótipo por PCR para seguir eGFP integração. 73% (n = 22) e 83% (n = 32) de embriões coletados tinha integrado o transgene para o CMV e a construção de Lentivirus de CAG, respectivamente (tabela 1). Embriões transgênicos foram então seccionado e imuno-manchado para eGFP. Conforme ilustrado na Figura 3, apenas eGFP dispersas células positivas são observadas com o promotor CMV (Figura 3, painel superior), Considerando que todas as células expressaram GFP quando utilizou-se o promotor CAG (Figura 3, painéis de média e inferior).
Com o promotor CAG, 96% dos embriões coletados de transgénicos ubiquitously expressa o transgene eGFP (tabela 1). Embora ambos os promotores são onipresentes, só o promotor CAG é capaz de expressão robusta de unidade do transgene em todas as células. Promotores onipresentes alternativos foram usados como fosfoglicerato quinase (PGK) e promotores do ubiquitin-C e produziu resultados semelhantes como os obtidos com o promotor CAG com baixos níveis de expressão de eGFP (dados não mostrados).
Na vivo mapeamento de regiões genômicas reguladoras para testar elementos de controle específicos do tecido.
Para um grande número de aplicações, expressão de transgenes de uma maneira específica de células em animais transgénicos é necessária. Além disso, a geração de animais transgênicos pode ser altamente instrumental para a capacidade de fragmentos de DNA genômicos regulamentares putativos para controlar a expressão específica de célula de um determinado gene de tela. Como exemplo, Lentivirus mediada por produção de animais transgénicos foi usada para mapear potenciadores de célula específica que controle de expressão de Neurogenin 3 (Neurog3)11. Neurog3 é um factor de transcrição do hélice-volta-hélice (bHLH) de base que controla o compromisso dos progenitores do pâncreas para o destino do sistema endócrino. Em Neurog3 ratos mutantes nulos, não há células endócrinas no pâncreas podem diferenciar19. Um fragmento de DNA de 2,2 kb localizado entre as posições-5284 e-3061 em relação ao Neurog3 transcrição iniciar site foi clonado em um vetor de Lentivirus montante que um promotor mínima da globina beta para expressão de unidade de um gene do repórter eGFP conforme descrito 11. uma construção de controle da mesma forma foi gerada por clonagem de um fragmento de 2,4 kb intergênicas localizado no cromossomo rato 6 (chr6: 14237279-14239685 em relação ao conjunto de genoma do rato mm9) na mesma espinha dorsal Lentivirus. Esta região genômica é localizada dentro de um 1 mega-base gene longo deserto entre genes Gpr85 e Ppp1r3a . Vetores de Lentivirus foram em seguida foram produzidos usando construções e nomeados Neurog3- enh-eGFP e Chr6-eGFP.
Ambos os vetores Lentivirus foram construídos e produziram (métodos complementares). Desde que não há células expressando Neurog3 estavam disponíveis atualmente, o título TU não pôde ser determinado. Alternativamente, o título foi medido como concentração de proteínas do capsídeo p24. Os 2 vetores foram injetados no espaço perivitelline de oócitos fertilizados e implantados em ratos fêmeas grávidos pseudo. Os embriões implantados foram coletados no dia embrionário 14,5 (E14.5), como nesta fase do desenvolvimento corresponde à expressão máxima da Neurog3 no pâncreas. Os embriões foram em seguida genótipo a seguir eGFP integração. 84% (n = 47) e 71% (n = 48) dos embriões coletados tinha integrado o transgene para Neurog3- enh-eGFP e Chr6-eGFP Lentivirus construções respectivamente (tabela 1). Para cada embrião, o broto pancreático foi dissecado e então seccionado para executar immunostaining. 92% de Neurog3- enh-eGFP embriões transgênicos expressa eGFP no pâncreas como ilustrado na Figura 4 painel superior (immunostaining representativo). Importante, a grande maioria dos eGFP positivo células também foram Neurog3 células expressando (Figura 4) indicando que o 2,2 kb Neurog3 enhancer é capaz de restringir a expressão eGFP dentro da população de células Neurog3 . Por oposição, nenhum dos embriões Chr6-eGFP expressa eGFP (painel inferior deFigura 4 e tabela 1) no pâncreas ou fora do pâncreas. Além disso, não observou-se nenhuma expressão ectópica de eGFP fora do pâncreas em Neurog3- enh-eGFP embriões11.
Para os 4 experimentos apresentados acima, uma descrição quantitativa precisa de cada etapa do processo é apresentada na tabela 1. Isto ilustra a eficácia global do processo. Com efeito, ao comparar os números de animais coletados que integrado o transgene com os números ovos fertilizados injetados, o rendimento global do processo é de 44% em média de. O mesmo rendimento com uma injeção de DNA pró-nuclear de uma construção contendo o realçador de Neurog3 fundido com a repórter da beta-galactosidase não exceda 3,1%.
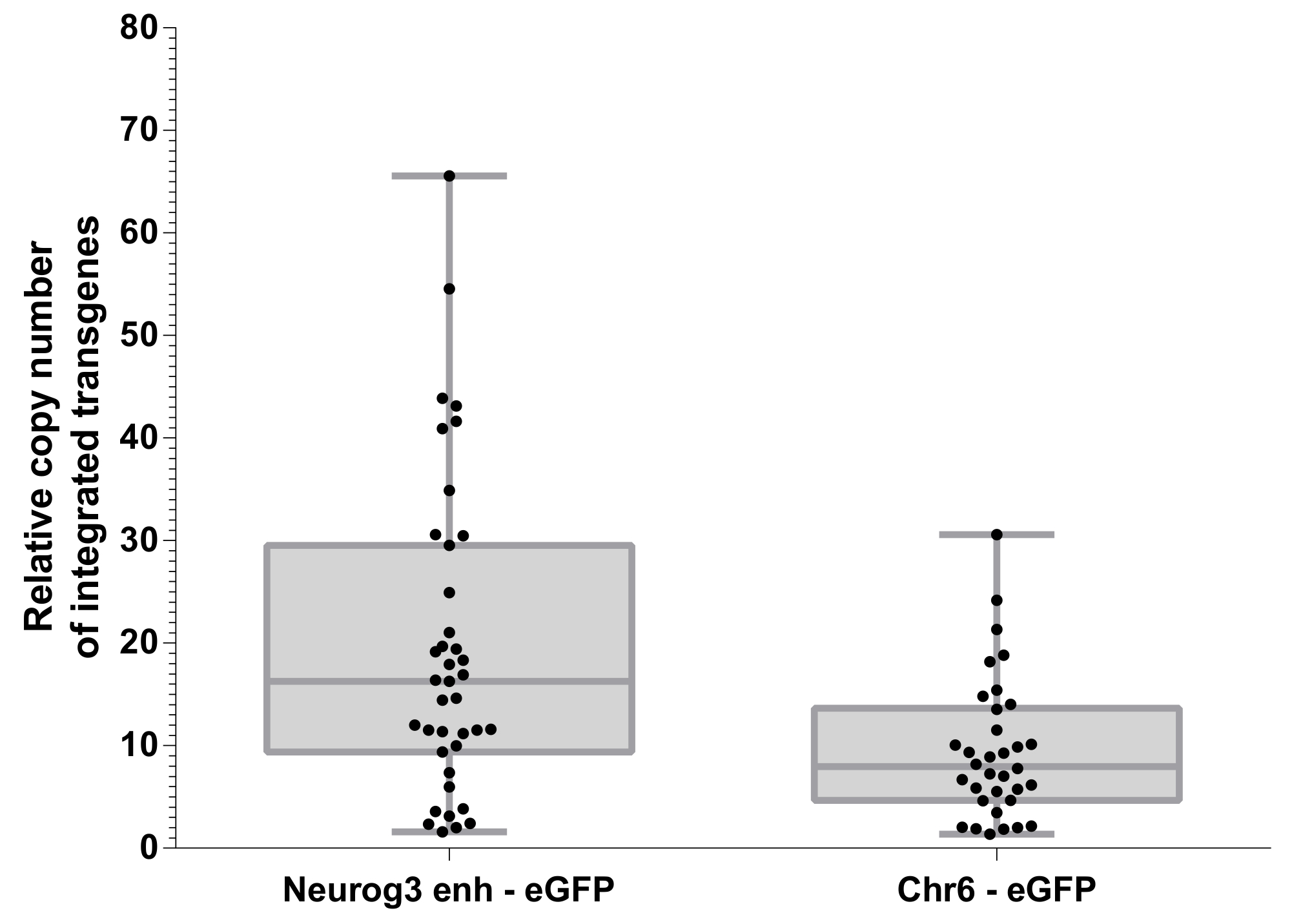
Transdução de oócitos fertilizados com um vetor de Lentivirus conduz à integração do transgene que pode ocorrer em vários sites de10,13. O número relativo de sites de integração do transgene foram avaliado usando PCR quantitativo no DNA genômico (Figura 5). Quantificação da eGFP integração foi determinada por PCR quantitativo (qPCR) e normalizada para gene Cdx2 que está presente em 2 cópias por genoma conforme descrito anteriormente,11. O número médio de sites de integração foi 19.36 ± 2.468 (no MEV mostrou) e 9.537 ± 1.186 (no MEV mostrou) em embriões gerados a partir de Neurog3- enh-eGFP e construção Chr6-eGFP, respectivamente. Curiosamente, os dois vetores Lentivirus usados para produzir estes animais apresentaram títulos virais diferentes. A concentração da proteína do capsídeo p24 eram de 124 ng / µ l para Neurog3- enh-eGFP vetor e de 52 ng / µ l para o controle de vetor de Chr6-eGFP. É mais provável que essa diferença de concentração serão responsáveis para a diferença significativa observada nos números do site de integração em ambos população de embriões transgênicos (Figura 5). Isto sugere que o número médio de integração de sites obtidos em um lote de ratos transgénicos fundador pode ser modulado usando estoques virais com títulos diferentes.
Importante, não observou-se nenhuma correlação direta entre a expressão de eGFP em Neurog3- enh-eGFP embriões transgênicos e o número de cópias do transgene que foram integrados. Em outras palavras, embriões que integrado simples ou múltiplas cópias do Neurog3- enh-eGFP transgene foram encontrados da mesma forma para eGFP expressa em células positivas Neurog3.

Figura 1: fluxograma do processo global Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Preparar a injeção micro pipetas e genotipagem.
(A) desenhos esquemáticos de microinjeção de pipetas para destacar as principais diferenças entre pipetas utilizadas para DNA ou vetor de Lentivirus injeções. Deixou o painel: a forma geral de ambos os tipos de pipeta é desenhada. O círculo tracejado realça a área ampliada da ponta da pipeta. Fotos das pontas de pipeta também são apresentadas. Observe que, para injeção de Lentivirus ponta precisa ser quebrado conforme indicado com a linha pontilhada e a imagem correspondente. Painel direito: exemplo de injeção de ovo definindo com a pipeta de exploração à esquerda, o óvulo fertilizado e a pipeta de injeção ou em um pronucleus ou no espaço perivitelline. Barras de escala = 50 µm. visualização (B) em gel de agarose dos produtos PCR eGFP amplificada do DNA genômico extraído de 8 diferentes embriões (faixa de 1 a 8). Apenas os embriões 1, 2, 3, 5, 6 e 8 tinham integrado o transgene eGFP. O DNA do plasmídeo pTrip utilizado para a produção de vetor de Lentivirus PGK-eGFP foi usado como controle positivo de PCR. Para o controle negativo, H2O substituído DNA na reação de PCR. MWM: marcador de peso molecular. BP = pares de base. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Promotores onipresentes conduzir a expressão do repórter eGFP em embriões transgênicos.
10 µm cryo-seções de embriões transgênicos foram coradas para visualizar a expressão eGFP (verde) e núcleos (azuis). Os embriões gerados com o promotor CMV Lentivirus construir (etiqueta superior esquerda) foram coletadas em E11.5. Os embriões gerados com a construção de Lentivirus CAG promotor (rótulo de esquerda inferior) foram coletados em E18.5. PB: broto pancreático, VSC: ventral da medula espinhal, Vt: vértebra, Li: fígado, Ms: músculo do cinto abdominal. Barras de escala = 50 µm clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Expressão específica de célula do gene repórter em embriões transgênicos é impulsionado pelo realçador de Neurog3 . 10 µm cryo-seções de E14.5 botões pancreáticas de embriões transgênicos foram coradas para visualizar a expressão de Neurog3 (vermelho), eGFP (verde) e núcleos (azuis), como descrito em métodos complementares). Neurog3 expressão está espalhado no pâncreas. Transgénicos embriões que integra o Neurog3- enh-eGFP construir eGFP expressa e a maioria de eGFP células positivas são Neurog3 positivo (painel superior). Os embriões gerados com a construção de Chr6-eGFP não estavam expressando eGFP (painel inferior). Barras de escala = 50 µm clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: número de cópia relativa de transgenes integrado. Quantificação dos sites de integração eGFP em relação ao gene CDX2 conforme descrito na seção de protocolo. Caixa plano de 25th para 75th percentil. Pontos representam os embriões transgênicos diferentes que foram gerados. A comparação de sites de transgenes integrado entre as duas construções de Lentivirus é significativamente diferente (teste t não pareado paramétrico, p = 0,001). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Tabela 1: relatório quantitativo passo a passo durante todo o procedimento. Durante o curso do procedimento, o número total de ovos ou embriões foram contado. A primeira coluna representa o número total de ovos que foram obtido os ovidutos das fêmeas superovuladas. Apenas ovos com clara 2 corpos polares e/ou visíveis pró-núcleos foram injetados e são relatados. Após a injeção e algumas horas em cultura foram implantados apenas injetados ovos que não foram lysed e tinham uma morfologia normal. Em seguida, o número total de embriões que foram coletados das fêmeas pseudopregnant é contado. Finalmente, os embriões que tinham integrado o transgene e expresso o repórter estão listados nas duas últimas colunas. Os mesmos recursos também são dadas para comparação com um experimento usando injeção de DNA pró-núcleos padrão. Aqui o transgene contido o realçador de Neurog3 dirigindo a expressão de um gene repórter de beta-galactosidase (Neurog3- enh-LacZ). Clique aqui para baixar este arquivo.
Discussão
A injeção de perivitelline de Lentivirus vetores em oócitos fertilizados descritos aqui resultou na produção de embriões transgênicos que rendeu mais de 70% dos embriões geneticamente modificados em relação ao número total de embriões coletados. Este resultado é consistente com relatórios anteriores e exemplifica a especificidade do procedimento2,7,10,11,12. Quando comparando todos os dados apresentados na tabela 1, podem ser realçadas características importantes. Em primeiro lugar, o número de ovos implantados correspondeu a todos ovos injetados tinham uma morfologia normal ou não foram lysed após algumas horas em cultura. 93% dos ovos injetados foram implantados sugerindo uma quase completa ausência de toxicidade rápida devido a injeção de um vetor de Lentivirus no espaço perivitelline. A situação é dramaticamente diferente quando se considera a injeção de DNA, desde que apenas 44% dos ovos injetados sobreviveram e foram implantados. Além disso, a proporção de embriões coletados em relação ao implantado ovos é idêntica entre os dois processos, sugerindo que não há toxicidade a longo prazo exacerbada de Lentivirus vetores. Em segundo lugar, quando o número de embriões que integrado o transgene em relação ao número de ovos injetados de expressar o rendimento global é mais de 10 vezes maior com injeção de vetor de Lentivirus comparada com injeção de DNA. Uma 86-fold diferença encontra-se mesmo quando se compara o número de embriões expressando transgene entre os dois processos usando a mesma construção de realçador de Neurog3.
Importante, rendimento de produção de transgénicos parece ser dependente do título de transdução dos vetores de Lentivirus usados. Em outras palavras, Lentivirus vetores produziram com um título acima 109 TU/mL são suficientes para obter tal alto rendimento. Conforme descrito na seção de protocolo, o volume injetado no espaço perivitelline está na faixa de 10 a 100 pl. Este volume representará particulas de Lentivirus ativas de 10 a 100. Em comparação com o padrão pró-núcleos injeções de DNA, o número total de animais fundador gerados com a mesma quantidade de animais nascidos é pelo menos 10-fold mais elevado quando usando vetores Lentivirus. Além disso, a penetrância de expressão do transgene é extremamente alta com este protocolo e observou-se ambos com onipresente e celulares promotores específicos, com exceção do promotor CMV. Por oposição aos promotores onipresentes celulares, o promotor CMV ativamente é fechado pela metilação de DNA20 e mostrou ser incapaz de manter a expressão de longo prazo sobre a transdução de células-tronco pluripotentes21. Isto poderia explicar o número muito limitado de eGFP expressando as células observadas nos embriões transgênicos. Portanto, vetores de Lentivirus são bem adaptadas para produzir animais transgénicos, em que a expressão de um transgene é controlada por um potenciador de célula específica. Importante, o protocolo pode ser usado para a tela para realçador atividade na vivo e encontrar mapa transcrição locais obrigatórios do fator dentro de regiões reguladoras11,12. Esta abordagem de triagem dificilmente pode ser executada usando o padrão transgênese. O número total de animais fundador precisava testar todas as construções diferentes e alcançar significância estatística exigiria dezenas de sessões de injeção, Considerando que pode ser obtido rapidamente com particulas de Lentivirus transgênese mediada.
Dentre as principais diferenças entre o procedimento padrão e o método com base Lentivirus reside na integração do transgene. Usando injeção pró-nuclear, transgenes integrar-se aleatoriamente como várias cópias em um único locus. Usando vetores Lentivirus, integração pode ocorrer em múltiplos loci (uma cópia por locus) sem ser estritamente aleatória. Clonando sites de integração usando Linear amplificação mediada por PCR (LAM-PCR), o grupo do Trono de D. demonstrou que transgenes integrar preferencialmente em regiões de cromatina aberta dos ovos fertilizados13. O viés de integração não deve interferir ou contribuir para a expressão do transgene nos ratos transgénicos. Integração durante Lentivirus transdução em um embrião de fase de uma célula ocorre na cromatina aberta que pode não estar ainda na configuração aberta mais tarde durante o desenvolvimento ou no adulto.
Além disso, ao analisar o número de cópias de transgenes integrado em animais de primeira geração (F0) ou embriões, é observada uma grande variação no número de transgene integrado. Neste estudo, verificou-se uma média de 19 cópias integradas com a construção de Neurog3-enh-eGFP. Este número de cópia grande poderia refletir altos níveis de mosaicismo. Sauvain et al realizaram um extenso estudo dos loci integrado em F0 animais gerados com o método mediado Lentivirus descrito aqui13. Eles seguiram 70 sites individuais de integração em 11 animais de F0 e examinaram as taxas de transmissão para cada site de F0 ratos transgênicos para seus descendentes de F1. A taxa global de transmissão de 44% para o transgene integrado individual sugere que eles foram mais frequentemente estabelecidos após a fase de S de embriões de uma célula, ou antes da fase S na fase de duas células. Com efeito, integração antes da fase S iria transmitir o transgene integrado para as duas células-filhas, enquanto integração após a fase de S que transmiti-los para apenas uma célula filha. Assim, o grau de mosaicismo para individuais transgenes integrado é mínimo em ratos transgénicos obtidos através desta técnica. Isso indica mais que mais integração ocorrerá dentro o primeiro h 12 correspondente para o tempo médio de produção da primeira clivagem das condições de cultura utilizado. Essa integração cinética é consistente com o descrito por lentivírus em de células linfoides T22.
Importante, com um elevado número de loci descobrindo transgenes integrada, que estabelece as linhas do mouse não seria razoável. O número de passagens para segregar todos estes loci seria consideravelmente alto. Isto representa uma importante limitação desse método que deve ser usado para triagem rápida dos efeitos do transgene ou para análise simultânea de múltiplos transgenes. No entanto, linhas de rato ainda podem ser estabelecidas, selecionando o animal F0 aqueles com o número de cópia menor do transgene integrado.
Desde a primeira descrição do pró-nuclear DNA injeção método1, melhorias foram feitas que contornar muitas das desvantagens do procedimento inicial. O primeiro conjunto de melhorias baseou-se na integração orientada em um locus preciso usando uma estratégia de troca de gaveta. Pró-nuclear injeção é executada usando qualquer CRE recombinases Flip ou PhiC31, juntamente com um fragmento de DNA integrativo ladeado com loxP, FRT ou attB sites, respectivamente. Nesta situação, o DNA integrativo é trocado com um fragmento integrado ladeado com a mesma recombinase site específico23,24. Apesar de até 60% dos animais de primeira geração pode ser transgênicos23 usando esse método, as limitações ligadas à tecnologia da injeção pró-nuclear ainda se aplicam. O segundo conjunto de melhoria é baseado em injeções citoplasmáticas de dois DNA circular, um carregando o fragmento para integrar e uma expressão que permite do qualquer Tol225, transposases de27 26 ou piggyBac bela adormecida. Usando esses métodos, são obtidos rendimentos elevados (> 30%), mas mais importante, a injeção citoplasmática é fácil de executar e contorna as restrições devido a injeção pró-nuclear como o protocolo baseado Lentivirus. Além disso, grandes fragmentos de DNA, tais como cromossomos artificiais bacterianos, podem ser integrados.
É claro que Lentivirus transgênese mediada não substituirá a norma nem os procedimentos melhorados. Ainda este método representa uma poderosa ferramenta para rápida modelo animal de produção e caracterização que reduz consideravelmente o tempo necessário para gerar o número adequado de animais com a variabilidade genética menos. Além disso, esta tecnologia pode ser aplicada diretamente para todas as cepas do mouse, incluindo quaisquer linhas transgénicas. Além disso, é importante mencionar que a paisagem global de geração de modelos animais romance está prestes a mudar com o desenvolvimento recente da tecnologia CRISPR/Cas9. Hoje, pró-nuclear injeções de proteína Cas9 juntamente com guia RNA permite a produção de genoma edição de modelos animais com uma eficácia de 40%28. Esta abordagem largamente poderia beneficiar do uso de Lentivirus transgênese mediada. Com efeito, o uso de vetores não-Integrativa de Lentivirus29 transitoriamente expressar ambos Cas9 e guia de RNAs pode resultar em ainda mais altos rendimentos de produção. A combinação das tecnologias mais recentes para produzir modelos animais relevantes e robustos beneficiaria mais internacionais grupos de pesquisa envolvidos no estudo da patogênese da doença e abordagens terapêuticas.
Divulgações
Os autores não têm nenhum conflito de interesses para divulgar.
Agradecimentos
Agradecemos a Magali Dumont e Rolando Meloni pela leitura crítica do manuscrito e iVector e Phenoparc ICM núcleos de assistência técnica na produção de vetor de Lentivirus e animal habitação respectivamente. Este trabalho foi apoiado do Translationnelles Institut Hospitalo Universitaire de Neurociências de Paris, o IHU-A-ICM, Investissements d'Avenir ANR-10-IAIHU-06. Relações públicas recebida financiamento para a associação de Langue Française pour l'Etude du Diabète et des Maladies Métaboliques (ALFEDIAM) e um conjunto JDRF / INSERM de concessão.
Materiais
| Name | Company | Catalog Number | Comments |
| PMSG 50UI | Sigma | G4527 | |
| hCG 5000UI | Sigma | CG5-1VL | |
| NaCl | Sigma | 7982 | |
| 100 mm petri dish | Dutsher | 353003 | |
| 4 wells Nunc dish | Dutsher | 56469 | IVF dish |
| M2 medium | Sigma | M7167 | |
| M16 medium | Sigma | M7292 | |
| 0,22 µm Syringe filter | Dutsher | 146611 | |
| Hyaluronidase Enzyme 30mg | Sigma | H4272 | mouse embryo tested |
| Insulin serynge | VWR | 613-3867 | Terumo Myjector |
| Curved forceps | Moria | 2183 | |
| Curved scissors | Moria | MC26 | |
| Aspirator tube assemblies for calibrated microcapillary pipettes | Sigma | A5177-5EA | |
| Borosilicate glass capillaries | Harvard apparatus | GC 100-10 | |
| Horizontal micropipette puller | Narishige | PN-30 | |
| Microforge | Narishige | MF-900 | |
| Inverted microscope | Nikon | Transferman NK2 5188 | Hoffman modulation contrast illumination is required |
| Micromanipulator | Eppendorf | Celltram air | |
| Controler of holding pipet | Eppendorf | Femtojet | |
| Mineral oil | Sigma | M8410 | mouse embryo tested |
| Microinjector | Eppendorf | Femtojet | Can be used to inject DNA or viral vectors |
| Dumont # 5 forceps | Moria | MC 40 | |
| vannas micro scissors | Moria | 9600 | |
| Isoflurane | centravet | ISO005 | ISO-VET 100% 250ml |
| ocrygel | centravet | OCR002 | |
| Povidone iodure | centravet | VET001 | vetedine 120ml |
| Buprenorphine | centravet | BUP002 | Buprecare 0,3Mg/ml 10ml |
| Tris-HCl | Sigma | T5941 | Trizma hydrochloride |
| EDTA | Sigma | E9884 | |
| SDS | Sigma | 436143 | |
| NaCl | Sigma | S7653 | powder |
| proteinase K | Sigma | P2308 | |
| oneTaq kit | NEB | M0480L | |
| Primers | Eurogentec | ||
| Strip of 8 PCR tube | 4titude | 4ti-0781 | |
| 96 well thermal cycler | Applied Biosystems | 4375786 | Veriti |
| Genomic DNA mini kit | invitrogen | K1820-02 | |
| Nanodrop 2000 | Thermo Scientific | ND-2000C | |
| qPCR Master mix | Roche | 4887352001 | SYBR Green |
| Multiwell plate 384 | Roche | 5217555001 | |
| qPCR instrument 384 well | Roche | 5015243001 | LightCycler 480 |
Referências
- Gordon, J. W., Scangos, G. A., Plotkin, D. J., Barbosa, J. A., Ruddle, F. H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proceedings of the National Academy of Science USA. 77 (12), 7380-7384 (1980).
- Bock, T. A., Orlic, D., Dunbar, C. E., Broxmeyer, H. E., Bodine, D. M. Improved engraftment of human hematopoietic cells in severe combined immunodeficient (SCID) mice carrying human cytokine transgenes. Journal of Experimental Medicine. 182 (6), 2037-2043 (1995).
- Miyakawa, Y., et al. Establishment of human granulocyte-macrophage colony stimulating factor producing transgenic SCID mice. British Journal of Haematology. 95 (3), 437-442 (1996).
- Hirabayashi, M., et al. A comparative study on the integration of exogenous DNA into mouse, rat, rabbit, and pig genomes. Experimental Animals. 50 (2), 125-131 (2001).
- Isola, L. M., Gordon, J. W. Transgenic animals: a new era in developmental biology and medicine. Biotechnology. 16, 3-20 (1991).
- Zufferey, R., Nagy, D., Mandel, R. J., Naldini, L., Trono, D. Multiply attenuated lentiviral vector achieves efficient gene delivery in vivo. Nature Biotechnology. 15 (9), 871-875 (1997).
- Lois, C., Hong, E. J., Pease, S., Brown, E. J., Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science. 295 (5556), 868-872 (2002).
- Ewerling, S., et al. Evaluation of laser-assisted lentiviral transgenesis in bovine. Transgenic Research. 15 (4), 447-454 (2006).
- Ritchie, W. A., Neil, C., King, T., Whitelaw, C. B. Transgenic embryos and mice produced from low titre lentiviral vectors. Transgenic Research. 16 (5), 661-664 (2007).
- Park, F. Lentiviral vectors: are they the future of animal transgenesis. Physiological Genomics. 31 (2), 159-173 (2007).
- van Arensbergen, J., et al. A distal intergenic region controls pancreatic endocrine differentiation by acting as a transcriptional enhancer and as a polycomb response element. PLoS One. 12 (2), e0171508 (2017).
- Friedli, M., et al. A systematic enhancer screen using lentivector transgenesis identifies conserved and non-conserved functional elements at the Olig1 and Olig2 locus. PLoS One. 5 (12), e15741 (2010).
- Sauvain, M. O., et al. Genotypic features of lentivirus transgenic mice. Journal of Virology. 82 (14), 7111-7119 (2008).
- Punzon, I., Criado, L. M., Serrano, A., Serrano, F., Bernad, A. Highly efficient lentiviral-mediated human cytokine transgenesis on the NOD/scid background. Blood. 103 (2), 580-582 (2004).
- Zennou, V., et al. The HIV-1 DNA flap stimulates HIV vector-mediated cell transduction in the brain. Nature Biotechnology. 19 (5), 446-450 (2001).
- Miyoshi, H., Blomer, U., Takahashi, M., Gage, F. H., Verma, I. M. Development of a self-inactivating lentivirus vector. Journal of Virology. 72 (10), 8150-8157 (1998).
- Yee, J. K., et al. A general method for the generation of high-titer, pantropic retroviral vectors: highly efficient infection of primary hepatocytes. Proceedings of the National Academy of Science USA. 91 (20), 9564-9568 (1994).
- Castaing, M., et al. Efficient restricted gene expression in beta cells by lentivirus-mediated gene transfer into pancreatic stem/progenitor cells. Diabetologia. 48 (4), 709-719 (2005).
- Gradwohl, G., Dierich, A., LeMeur, M., Guillemot, F. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proceedings of the National Academy of Science USA. 97 (4), 1607-1611 (2000).
- Scharfmann, R., Axelrod, J. H., Verma, I. M. Long-term in vivo expression of retrovirus-mediated gene transfer in mouse fibroblast implants. Proceedings of the National Academy of Science USA. 88 (11), 4626-4630 (1991).
- Norrman, K., et al. Quantitative comparison of constitutive promoters in human ES cells. PLoS One. 5 (8), e12413 (2010).
- Vandegraaff, N., Kumar, R., Burrell, C. J., Li, P. Kinetics of human immunodeficiency virus type 1 (HIV) DNA integration in acutely infected cells as determined using a novel assay for detection of integrated HIV DNA. Journal of Virology. 75 (22), 11253-11260 (2001).
- Ohtsuka, M., et al. One-step generation of multiple transgenic mouse lines using an improved Pronuclear Injection-based Targeted Transgenesis (i-PITT). BMC Genomics. 16, 274 (2015).
- Tasic, B., et al. Site-specific integrase-mediated transgenesis in mice via pronuclear injection. Proceedings of the National Academy of Science USA. 108 (19), 7902-7907 (2011).
- Sumiyama, K., Kawakami, K., Yagita, K. A simple and highly efficient transgenesis method in mice with the Tol2 transposon system and cytoplasmic microinjection. Genomics. 95 (5), 306-311 (2010).
- Garrels, W., et al. Cytoplasmic injection of murine zygotes with Sleeping Beauty transposon plasmids and minicircles results in the efficient generation of germline transgenic mice. Biotechnology Journal. 11 (1), 178-184 (2016).
- Ding, S., et al. Efficient transposition of the piggyBac (PB) transposon in mammalian cells and mice. Cell. 122 (3), 473-483 (2005).
- Aida, T., et al. Cloning-free CRISPR/Cas system facilitates functional cassette knock-in in mice. Genome Biology. 16, (2015).
- Philippe, S., et al. Lentiviral vectors with a defective integrase allow efficient and sustained transgene expression in vitro and in vivo. Proceedings of the National Academy of Science USA. 103 (47), 17684-17689 (2006).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados