
Method Article
Imagerie de Ca2 + réponses au cours de l’Infection des cellules épithéliales de Shigella
Dans cet article
Résumé
Nous présentons ici les protocoles afin de visualiser les réponses de calcium (Ca2 +) induites par les cellules HeLa infectées par Shigella. En optimisant les paramètres de l’infection bactérienne et d’imagerie avec Ca2 + des sondes fluorescentes, atypique globales et locales Ca2 + signaux induits par des bactéries sur une gamme étendue de la cinétique de l’infection sont caractérisés.
Résumé
Ca2 + est un ion ubiquitaire impliqué dans tous les processus cellulaires connus. Alors que Ca2 + réponses globales peuvent affecter le sort de la cellule, les variations locales de Ca2 + cytosolique des concentrations libre, liées à la libération de réserves internes ou un influx à travers les canaux de la membrane plasmique, régulent les processus de cellules corticales. Pathogènes qui respecte ou envahir l’hôte cellules déclencheur une réorganisation du cytosquelette d’actine qui sous-tendent la membrane plasmique hôte, affectant probablement Ca2 + signalisation tant mondial que local. Parce que ces événements peuvent se produire dans les basses fréquences de façon Pseudo-aléatoire stochastique plus longue cinétique, l’analyse des signaux de Ca2 + induite par des agents pathogènes soulève des défis techniques majeurs qui doivent être abordées.
Nous rapportons ici, protocoles pour la détection de Ca2 + des signaux les et mondiaux sur une infection Shigella des cellules épithéliales. Dans ces protocoles, artefacts liés à une exposition prolongée et photovieillissement associée à l’excitation de Ca2 + des sondes fluorescentes sont dépannez-effectué en contrôlant rigoureusement les paramètres d’acquisition au cours de périodes définies au cours d’une Shigella invasion. Des procédures sont mises en œuvre pour analyser rigoureusement l’amplitude et la fréquence des signaux2 + Ca cytosoliques globales au cours de la cinétique d’infection étendue à l’aide de la sonde chimique Fluo-4.
Introduction
Ca2 + réglemente tous les processus cellulaires connus, notamment la réorganisation du cytosquelette, les réponses inflammatoires et voies de mort cellulaire, associés à des interactions hôte-pathogène1,2,3. Dans des conditions physiologiques, des concentrations2 + Ca cytosoliques basales sont faibles, dans les centaines de gamme nM, mais peuvent être soumises à des augmentations transitoires lors de la stimulation agoniste. Ces variations montrent souvent comportement oscillatoire sous l’action des pompes et des canaux à la membrane de plasma et le réticulum endoplasmique. Le modèle de ces oscillations est caractérisé par la période, la durée et amplitude de Ca2 + augmente et est déchiffré par les cellules qui, à leur tour, déclenchent des réponses spécifiques dans ce qu’on appelle le Ca2 + code4,5 . Une augmentation soutenue de la concentration de Ca2 + cytosolique dans des conditions pathologiques peut-être conduire à la mort de cellule associée à la perméabilisation des membranes mitochondriales et le déblocage de pro-apoptotiques et nécrotiques facteurs6, 7.
Shigella, l’agent causal de la dysenterie bacillaire, envahit les cellules épithéliales en injectant des effecteurs dans les cellules de l’hôte à l’aide d’un type III sécrétion système (T3SS)8,9. Ca2 + signaux les et mondiaux provoquées par la T3SS est associé liée une invasion de Shigella des cellules hôtes. En ce qui concerne le pore-forming toxines, le translocon T3SS qui insère dans les membranes des cellules hôtes et est nécessaire pour l’injection des effecteurs T3SS est probablement responsable de l’activation de PLC et l’inositol (1, 4, 5) trisphosphate (InsP3)-dépendant Ca2 + Communiqué. Communique de la combinaison de la stimulation localisée de PLC et l’accumulation de l’actine polymérisée dans les endroits d’un résultat d’invasion de Shigella dans un anormalement longue durée dépendant InsP3 Ca2 + 10. Le type III processeur d’effets IpgD, une phosphatidyl 4,5 bisphosphate (PIP2) -4-phosphatase, limite l’espace local de PIP2, contrôlant ainsi la quantité de substrat disponible pour PLC générer des InsP3, ce qui contribue à l’isolement des locaux Ca2 + réponses à l’invasion bactérienne sites11,12. Ces locaux Ca2 + réponses susceptibles de contribuer à la polymérisation de l’actine à Shigella invasion sites10. Réponses2 + Ca globales qui sont aussi provoquées par Shigella, cependant, sont indispensables pour le processus d’invasion bactérienne mais déclencher l’ouverture de connexine hémicanaux à la membrane plasmique et la libération d’ATP dans l’extracellulaire compartiment. ATP libérée, agissant de manière paracrine, stimule à son tour, Ca2 + oscillatoires réponses dans des cellules à côté de la cellule infectée. IpgD est également responsable de la mise en forme Ca2 + réponses globales dans des réponses isolées erratiques avec la dynamique lente. Finalement, après une infection bactérienne prolongée, IpgD conduit à l’inhibition des signaux2 + Ca induite par l’InsP3. Par le biais de son interférence avec la signalisation de Ca2 + , IpgD retarde un Ca2 +-activation dépendante calpaïne permettant le démontage des structures d’adhérence focale et le détachement prématuré d’infecté les cellules13.
Alors que les signaux de Ca2 + sont impliqués dans les aspects critiques de la pathogenèse, l’utilisation d’un micro-organisme soulève un certain nombre de défis techniques qui ne sont pas rencontrés dans les études classiques agoniste. Protocoles décrits ici utilisent le couramment utilisé Ca2 + chimique indicateur fluorescent Fluo-4 que nous avons conçu pour caractériser les signaux locaux de Ca2 + au cours d’une infection de Shigella . Étapes essentielles pour la détection de ces signaux sont discutés, ainsi que les procédures mises en place pour leur analyse quantitative qui sert à caractériser le rôle des effecteurs bactériennes dans la signalisation de Ca2 + .
Protocole
1. les préparatifs
-
Préparation de bactéries
- Les bactéries de la plaque — souche sauvage deShigella exprimant l’adhésine aloudi (M90T-aloudi) — sur un trypticase agar de soja (TCS) plaque contenant 0,01 % du rouge Congo (CR) et les incuber pendant 18 heures à 37 ° C.
Remarque : Pour augmenter leur reproductibilité, les plaques de Shigella , obtenus à l’étape 1.1.1 sont stockés à 4 ° C et doivent être utilisés dans une semaine, parce que toutes les colonies sur milieu CR finalement seront allume en rouges au fil du temps. - Inoculer TCS bouillon pré-culture en choisissant 3 colonies rouges des plaques strie.
Remarque : L’inoculation avec 3 colonies est effectuée afin de limiter la probabilité de choisir un seul clone dont plasmides de virulence a fait l’objet d’une recombinaison génétique. - Cultiver des cultures bactériennes liquides dans un incubateur à agitation pendant 16 h à 37 ° C à 200 tr/min. Ajouter ampicilline à une concentration finale de 75 mg/mL. Concentrations d’antibiotique ne doivent pas dépasser 3 x la concentration minimale inhibitrice, car un excès d’antibiotique peut conduire à la perte du plasmide grande virulence.
Remarque : Le maintien du plasmide Shigella virulence doit être vérifié en ensemençant les cultures bactériennes sur des plaques de CR additionnés de concentrations d’antibiotique appropriées. La présence de colonies blanches indique perte de plasmides. - Ensemencer la culture TCS avec la culture bactérienne préalable à une dilution au 1/100. Il incuber à 37 ° C dans un incubateur à agitation pendant 2 h à 200 tr/min. Veiller à ce que la densité optique à 600 nm (OD600nm) est de 0,2 - 0,4.
- Centrifuger la culture bactérienne pendant 2 min à 13 000 x g à 21 ° C et Resuspendre le culot dans un volume équivalent de EM milieu (120 mM NaCl, KCl 7 mM, 1,8 mM CaCl2, 0,8 mM MgCl2, glucose 5 mM et 25 mM HEPES pH = 7,3).
- Diluer la suspension bactérienne dans un tampon EM à une finale de OD600nm de 0,1 et utiliser immédiatement ou stockez-le à 21 ° C et utiliser dans le prochaine 60 min.
- Les bactéries de la plaque — souche sauvage deShigella exprimant l’adhésine aloudi (M90T-aloudi) — sur un trypticase agar de soja (TCS) plaque contenant 0,01 % du rouge Congo (CR) et les incuber pendant 18 heures à 37 ° C.
-
Préparation de cellules
- Les cellules HeLa culture en DMEM avec 1 g/L de glucose additionné de 10 % de sérum de veau foetal (FCS) et cultivent à 37 ° C, avec 10 % de CO2. Croître TC-7 grandi DMEM avec 4,5 g/L de glucose, contenant 10 % FCS et les acides aminés non essentiels dans un incubateur à 37 ° C contenant 10 % de CO2.
- Pour l’entretien de la cellule, fractionner les cellules régulièrement afin d’éviter une inhibition de la croissance-contact liée aux États confluentes ; Cela est essentiel pour un Ca2 + sonde chargement/transfection à rendement élevé.
- Plaque de cellules HeLa sur lamelles circulaires stériles 25 mm de diamètre en plaques 6 puits à une densité de 3 x 105 cellules/puits la veille de l’expérience pour les cellules HeLa.
- Pour les TC-7 cellules, trypsinize et compter les cellules. Les semences à 4 x 105 cellules/puits et incuber les 5-7 jours à 37 ° C dans un 10 % de CO2-incubateur avant les expériences afin de permettre une polarisation.
- Actualiser le milieu de culture cellulaire de tous les jours jusqu’au jour de l’expérience.
Remarque : Fond en verre diamètre 35 mm jetable chambres peut-être également être utilisé comme une alternative aux puits contenant des lamelles couvre-objet. Si ces derniers sont utilisés, le contrôle de la température grâce à une platine chauffante ou objectif n’est pas suffisant, et une chambre d’incubation thermostat monté sur la platine du microscope est nécessaire.
- Actualiser le milieu de culture cellulaire de tous les jours jusqu’au jour de l’expérience.
- Le jour de l’expérience, retirez le support et laver les cellules 3 x avec 1 mL EM médium à 21 ° C.
- Charger les cellules avec 3 mM Fluo-4-AM14 un tampon EM pendant 30 minutes à 21 ° C.
Remarque : Le chargement de cellules HeLa Sub confluentes ne soulève pas de problèmes majeurs, le chargement des cellules polarisées de TC-7 est souvent hétérogène et peu efficace. Pour ces cellules, nous avons trouvé que l’efficacité de chargement est renforcée par l’ajout de l’agent de dispersion — acide Pluronique — à une concentration finale de 0,1 % et de l’anion-transporteur inhibiteur bromosulfophtalein à une concentration finale de 20 µM à la Contenant fluo-4-AM EM. L’utilisation de produits chimique ratiométrique sondes ou codé génétiquement-FRET sondes nécessitant une double-acquisition n’est pas compatible avec la vitesse d’acquisition requise pour l’imagerie des réponses locales du Ca2 + . - Laver les échantillons 3 x avec EM mettre en mémoire tampon et plus loin les incuber dans 1 mL de tampon EM à 21 ° C, permettant l’hydrolyse de la portion de l’AM. Utilisation Fluo-4 cellules chargées immédiatement ou dans les prochain 90 min (maintenue à 21 ° C).
2. infection et Acquisition d’images des réponses locales Ca2 +
- Placer la lamelle contenant les cellules chargées de Fluo-4 dans une chambre d’imagerie ou de la chambre de fond en verre. Laver les échantillons dans la salle d’imagerie ou jetables de fond en verre chambre 3 x avec tampon EM pour éliminer les composés potentiellement résultant de la lyse cellulaire. Ajouter 1 mL de tampon d’EM.
- Placer la chambre sur une scène de microscope de fluorescence inversé chauffée à 33 ° C.
Remarque : Cette température est sélectionnée comme un compromis entre les températures optimales compatibles avec le processus d’invasion de Shigella et le ralentissement des réponses du Ca2 + pour visualiser les réponses locales. Nous utilisons un 63 X objectif à immersion d’huile (NA = 1,25) équipé d’anneaux de contraste de phase. - Sélectionnez un champ de microscopie et de définir les paramètres d’acquisition, y compris les temps d’exposition et binning si nécessaire, pour optimiser le signal fluorescent Fluo-4.
Remarque : Les grands enjeux du local Ca2 + imagerie sont la faible amplitude et la faible durée des réponses, nécessitant une acquisition au moins tous les 30 m plusieurs disponibles dans le commerce rétro-éclairé EM-CCD ou C-MOS caméras présentent le nécessaire sensibilité pour détecter Ca2 + réponses locales à l’aide de Fluo-4. Sensibilité de détection est également une question importante pour limiter le photovieillissement ou objets tels que des réponses spontanées, liées à l’excitation fluorescente Fluo-4 à haute fréquence. Ici, un éclairage à base de LED de 470 nm, avec un filtre passe-bande d’excitation de 480 ± 40 nm, un filtre dichroïque 505 nm et une émission de passe-bande nm ± 30 527 utilisé à 5 % de son intensité maximale combinée avec un filtre de densité neutre 1,0 ont été utilisés afin de minimiser ces problèmes sous un courant d’acquisitions des durées limitées. Une analyse du Ca2 + signaux locaux de lots de 120 s flux s’est déroulée régulièrement. Dans ces conditions de faible éclairage, fluorophore photoblanchiment n’a pas été observé au cours de l’acquisition de min-stream 2. Les acquisitions successives sur le même échantillon peuvent être effectuées, autant de domaines différents sont utilisés. En règle générale, un premier flux d’acquisition est effectué sur un champ de contrôle en l’absence de stimulation de bactéries pour s’assurer de l’absence de réponses spontanées. Si on observe des réponses spontanées, les échantillons sont lavés avec un tampon EM jusqu'à ce qu’on observe aucune réponse. - Pour ajouter des bactéries à l’échantillon, retirer 500 µL de tampon de l’EM de la chambre et ajouter 500 µL de la suspension bactérienne préparée à l’étape 1.1.6 pour obtenir une finale de OD600nm de 0,05, avec les soins appropriés afin d’éviter de déplacer le champ sélectionné de la microscopie ; les volumes s’assurer un bon mélange de bactéries et une répartition homogène de la bactérie sur les échantillons de cellules.
- Effectuer une acquisition suite à l’ajout de bactéries. Vous pouvez également effectuer une acquisition 10 min après l’ajout de bactéries, qui correspond au temps nécessaire pour que les bactéries dans les sédiments sur les cellules dans les conditions utilisées. À la fin du flux de l’acquisition, acquérir une image à contraste de phase du champ sélectionné pour visualiser les bactéries communiquant avec les cellules et les volants de membrane associées aux sites d’invasion bactérienne.
- Répétez la procédure d’acquisition comme au point 2.5, à des intervalles qui sont prêtent à la durée de l’acquisitions d’image, fichiers d’épargne et sélection d’un nouveau champ à couvrir l’ensemble du processus.
Remarque : Pour les Shigella, nous avons trouvé que l’acquisition diffuse toutes les 5 min pendant 20 min, après la sédimentation bactérienne de 10 min, permise de couvrir la majorité des événements invasion. - Ajouter à la fin de la procédure d’acquisition, une concentration finale de 2 µM de Ca2 + ionophore ionomycine échantillons pour déterminer l’amplitude maximale des signaux Ca2 + . Acquisition d’images toutes les 3-5 s jusqu'à stabilisation du signal, généralement pour moins de 10 min.
- Suivre en ajoutant Ca2 + chélateur EGTA à une concentration finale de 10 mM pour déterminer le signal de fluorescence en l’absence de Ca2 +. Acquisition d’images toutes les 3-5 s jusqu'à stabilisation du signal, généralement pour moins de 10 min.
Remarque : En raison de la dynamique relativement lente des Ca2 + variations globales, effectuer ces acquisitions toutes les 3-5 s (pas en mode streaming), permettant de Ca2 + imagerie sur le même champ microscopique durant les traitements successifs.
3. analyse
- Exprimer des variations cytosoliques de Ca2 + au fil du temps comme le pourcentage de ΔF/F0, où ΔF représente la variation de l’intensité de fluorescence moyenne de la région d’intérêt (ROI) et F0 la fluorescence de base dans le même ROI, auquel une région non pertinents du champ dépourvu de cellules est soustrait.
- Supprimer les niveaux de fluorescence basale pour chaque cellule dans différents domaines, correspondant aux niveaux de référence ; ces niveaux peut être déterminé sans ambiguïté en raison de la faible fréquence et de relativement courte durée de Ca2 + réponses locales dans une acquisition de flux donné.
Remarque : Quantifier les caractéristiques frappantes des réponses liées aux questions posées et le modèle pathogène étudiés. Classiquement, différents paramètres sont pris en compte pour analyser les signaux Ca2 + , y compris la fréquence des cellules montrant locales ou globales Ca2 + réponses, ainsi que la durée, amplitude et la fréquence de ces réponses. Dans le cas de Shigella et les réponses locales, un autre aspect important de l’analyse est l’association spatiale des réponses avec des sites d’invasion bactérienne.
- Supprimer les niveaux de fluorescence basale pour chaque cellule dans différents domaines, correspondant aux niveaux de référence ; ces niveaux peut être déterminé sans ambiguïté en raison de la faible fréquence et de relativement courte durée de Ca2 + réponses locales dans une acquisition de flux donné.
- Afin de quantifier le pourcentage de cellules sensibles présentant des réponses globales ou locales, attirer ROIs dans les cellules, correspondant aux variations d’intensité de Fluo-4 séries.
- La valeur des paramètres stricts pour identifier le Ca2 + les réponses locales, telles que l’amplitude des variations de fond ci-dessus triple du niveau de référence cellule fluorescence moyenne intensité ou une durée d’au moins 200 ms, pour éviter de marquer un faux positif.
Remarque : En règle générale, même de petites Ca2 + réponses locales se distingués du bruit de fond par leur profil. La ROIs doit être suffisamment grande pour intégrer assez signaux fluorescents pour permettre la détection des variations locales. Selon l’intensité de la détection de Fluo-4, ces ROIs peuvent correspondre aux cercles d’un diamètre allant de 2 à 5 mM. - Mesurer les variations de l’intensité de fluorescence Fluo-4 moyenne dans 2 régions dans une zone distincte de la même cellule, afin de déterminer si les réponses sont globales ou locales.
Remarque : L’analyse d’un grand nombre de cellules est facilitée par l’utilisation de l’imagerie des logiciels permettant l’affichage de l’écran en direct des variations de l’intensité de la fluorescence moyenne au fil du temps pour les régions sélectionnées. Les logiciels d’imagerie disponibles dans le commerce a une telle option, mais cela peut aussi être effectuée en utilisant le logiciel gratuit Icy , en utilisant le plugin ROI intensité Evolution . - Déterminer le pourcentage de cellules présentant des réponses locales.
Remarque : Ce pourcentage est calculé à partir du nombre total de cellules analysées dans le domaine de la microscopie, qui doit être en nombre suffisant pour soutenir un statistiquement l’importance à l’aide d’un test non paramétrique. - Déterminer la durée des réponses locales.
Remarque : Réponses locales Ca2 + peuvent être davantage distingués selon leurs gammes de durée (c'est-à-dire, moins de 500 ms, entre 5 et 10 s, ou supérieure à 10 s) et leur association avec des sites d’invasion de Shigella .
- La valeur des paramètres stricts pour identifier le Ca2 + les réponses locales, telles que l’amplitude des variations de fond ci-dessus triple du niveau de référence cellule fluorescence moyenne intensité ou une durée d’au moins 200 ms, pour éviter de marquer un faux positif.
- Quantifier l’amplitude des réponses. Tout d’abord, calibrer le Ca2 + et cellule fond niveau maximal déterminé comme au point 2.1.7. Puisque la charge Fluo-4 peut varier pour chaque cellule, effectuer ces déterminations pour des cellules individuelles dans le domaine.
- Exprimer l’amplitude des réponses sous forme d’un pourcentage relatif de la réponse maximale.
- Effectuer des tests statistiques à l’aide d’un test de normalité, suivi d’un test paramétrique pour analyser les différences potentielles de l’amplitude des réponses entre les échantillons.
- Déterminer l’association entre ces réponses par rapport à des sites d’invasion bactérienne par leur localisation respective pour les volants de membrane induite par la bactérie visualisées dans les images de contraste de phase correspondants.
Résultats
Invasion de Shigella est associée à atypique durable Ca2 + réponses locales :
À la suite du protocole mentionné ci-dessus, les cellules HeLa Fluo-4-chargés ont été contestées avec WT Shigella et acquisitions de flux ont été effectuées pour analyser les signaux de Ca2 + . Une expérience représentative est illustrée à la Figure 1, avec une série d’images de Time-lapse de l’intensité de la fluorescence de la sonde Fluo-4 en moyenne dans une région d’intérêt pour une seule cellule et l’image correspondante de contraste de phase (Figure 1 a, à gauche Groupe d’experts). Le site d’invasion de Shigella se caractérise par la membrane volants détectés à l’image de contraste de phase (Figure 1 a, flèche). Local atypique augmente en gratuit Ca cytosolique2 + sont observées sur le site d’invasion de Shigella avec une amplitude variable et des durées allant de 2,5 à 5 s (Figures 1 a et 1 b, flèches), suivie par une augmentation globale des patients infectés cellulaire (Figure 1 b, pointes de flèches).
Le type III processeur d’effets IpgD régit la transition du local au global Ca2 + réponses induites par Shigella dans les cellules HeLa :
L’analyse des signaux de Ca2 + induite par le sauvage Shigella et en une souche mutante isogéniques pour le type III processeur d’effets IpgD, une phosphatidyl 4, 5 bisphosphate phosphatase, ont indiqué que cette dernière souche induisait plus globale et moins des réponses locales atypiques avec une longue durée (RATPs) avec 11,3 ± 4.4 (SEM) % et 40,3 ± 7,5 (SEM) % des cellules sensibles avec des réponses globales dans le cas d’un mutant WT et ipgD, respectivement (Figure 1)15. Ce mutant ipgD obtient des réponses locales à une fréquence similaire que la souche WT.
Shigella inhibent Ca global InsP3-dépendante2 + augmente dans les cellules HeLa lors cinétique infection prolongée :
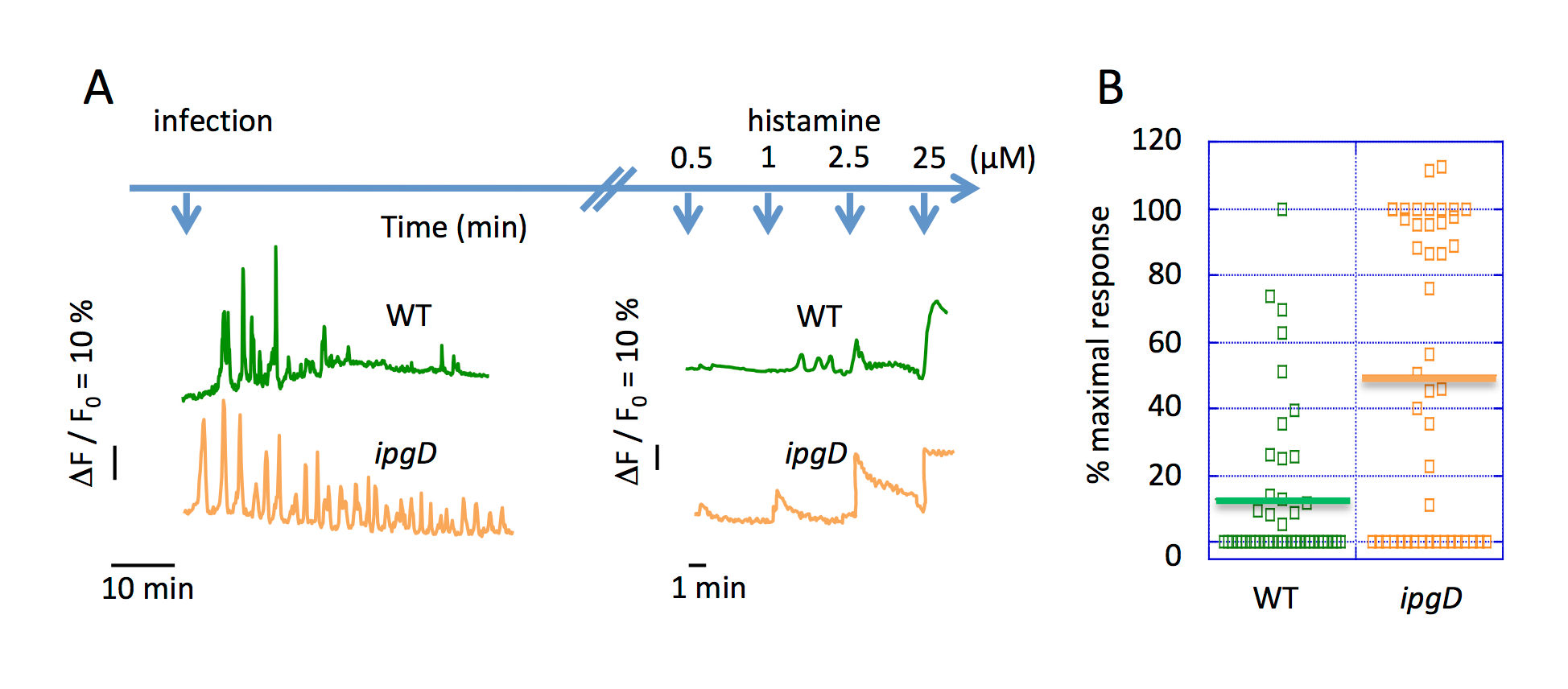
L’imagerie de Ca2 + réponses globales sur la cinétique de l’étendue de l’infection a signalé une diminution de la fréquence des réponses 30 min après le défi avec sauvage de Shigella (Figure 2 a). A observé une diminution similaire dans la fréquence des réponses du Ca2 + dans les TC-7 cellules infectées pendant 30 min avec sauvage Shigella15. La quantification des lymphocytes dose-dépendante de l’histamine Ca2 + agoniste illustre l’inhibition d’une libération2 + Ca InsP3-dépendante aux derniers stades d’une infection à Shigella . WT Shigella mène à atypique isolé Ca2 + réponses durant les premières 30 min de l’infection et après encore l’incubation, une diminution drastique de l’amplitude et la fréquence des réponses2 + Ca a été observée (Figure 2 b).

Figure 1 . Réponses locales et mondiales Ca2 + au cours de l’invasion de Shigella . Les cellules HeLa ont été chargées avec Fluo-4-AM et contestés avec WT Shigella. (A), le panneau de gauche montre des images à contraste de phase. Le panneau de droite montre la série temporelle de l’intensité moyenne de fluorescence Fluo-4 avec le code couleur représenté sur la droite. Le temps depuis le début de l’acquisition est indiqué en secondes, 10 min après le défi bactérien. Les flèches indiquent les foyers de l’invasion. Les cercles décrits correspondent aux régions analysées en (B) où les traces avec la couleur correspondante représentent les variations dans l’intensité de fluorescence moyenne de Fluo-4 sur la ligne de base. Les flèches indiquent les réponses locales. Les flèches indiquent les réponses globales. (C) il s’agit d’un schéma illustrant la détermination de la durée des réponses du Ca2 + . La ligne pointillée horizontale indique le taux basal Ca2 + (F,0) ; les barres verticales indiquent les variations de fond. La ligne verticale en pointillés indique l’amplitude maximale de la réponse ; les barres horizontales indiquent la durée de la réponse à l’amplitude maximale. (D), c’est le pourcentage de cellules sensibles ± SEM montrant les réponses locales et Ca2 + réponses globales induite par 5 min suivant l’infection avec le WT Shigella (barres gris foncé) ou le mutant ipgD (barres gris clair). RATPs sont parmi les réponses Ca2 + locales qui durent de plus de 5 s. N = 4 ; > 60 cellules pour chaque fois. Test de Wilcoxon, *P < 0,05 ; P < 0,01. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 . Inhibition des réponses2 + Ca globales au cours de la longue cinétique d’infection de Shigella . (A), le bleu haut flèches représentent l’échelle de temps du défi bactérien et l’histamine à des concentrations indiquées. Ce panneau montre les traces représentatives de cellule unique Ca2 + variations globales suite à l’infection par le WT Shigella (vert) et ipgD souche mutante (orange). (B) ce panneau indique le pourcentage de l’amplitude des réponses Ca2 + par rapport à la réponse maximale, lors d’une stimulation à l’histamine μM 0,5 des cellules infectées par des souches indiquées pendant 90 min. La barre horizontale solide représente le pourcentage maximal moyen. Test de Wilcoxon, **P < 0,01. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Discussion
Cet article décrit le protocole que nous avons conçu pour suivre les signaux locaux de Ca2 + pendant la cinétique relativement courte d’une invasion de Shigella , en plus de Ca2 + réponses globales au cours de la longue cinétique de Shigella. Ci-dessous, la clé se trouvent des questions qui doivent être abordées afin d’optimiser la détection de Ca2 + signaux tout en minimisant toute ingérence dans les processus biologiques.
Chimiques vs codé génétiquement Ca2 + sondes :
Pour Ca2 + variations locales de l’image, nous avons utilisé la sonde chimique Fluo-4 en raison de son rendement quantique élevée et sa dynamique de réponse rapide. La vitesse requise pour l’acquisition exclut également l’utilisation de ratiométrique sondes, puisqu’une acquisition double longueur d’onde ne peut pas être effectuée. L’utilisation de micro-organismes pathogènes, cependant, peut-être interférer avec les procédures standard à l’aide de sondes chimiques Ca2 + . Par exemple, une infection de la cellule prolongée pendant 60 min par Shigella empêche n’importe quelle cellule de chargement avec sondes chimiques, probablement à cause des altérations induites par le pathogène des membranes plasmiques cellule hôte. Régulièrement, une diminution de la fluorescence de Fluo-4 à la cellule est observée au cours de la cinétique de l’infection à long, en raison de la perte cytoplasmique de cette sonde. Par conséquent, la mise en œuvre d’un contrôle tel que l’ajout de l’ionomycine pour déterminer la fluorescence aux concentrations de Ca2 + maximales à la fin des acquisitions est critique pour identifier les cellules sensibles. En outre, polarisé les cellules épithéliales intestinales pertinentes pour une infection par des pathogènes semblent réfractaires au chargement de la sonde de Ca2 + et nécessitent des procédures spécifiques de chargement. Alors que l’utilisation d’un codé génétiquement Ca2 + reporter (GECR) peut aider à résoudre certaines de ces questions, il présente également d’autres défis. L’efficacité de transfection dans les systèmes de cellule hôte peut représenter une limitation sérieuse, surtout si elle se superpose avec un faible rendement infectieux. En outre, les caractéristiques d’une sensibilité aux variations de Ca2 + de la plupart des GECRs ne sont pas compatibles avec la cinétique rapide requise pour l’analyse locale de Ca2 + . Enfin, l’expression de la GECR peut interférer avec processus médiés par les agents pathogènes. Nous ne discuterons pas l’utilisation du GECR pour l’étude du Ca2 + signalisation lors d’une interaction hôte-pathogène. L’ingénierie récente et en cours des différents « réponse rapide » GECR mériterait probablement chercheurs pour revenir sur leur utilisation dans de futures études.
Calendrier d’acquisition de Ca2 + réponses locales au cours de l’infection bactérienne :
L’imagerie des réponses locales du Ca2 + nécessite une vitesse élevée d’acquisition d’images impliquant une excitation de fluorescence soutenue de la sonde Fluo-4. Cause le photovieillissement lié à l’acquisition de haute fréquence, il est essentiel que l’intensité de l’excitation de fluorophore léger est maintenue au minimum requis pour obtenir un rapport signal-bruit suffisant avec un temps de pose n’excédant ne pas 30 m, nous avons eu bons résultats en utilisant un système de LED à 5 % de son intensité maximale, combiné avec un filtre de densité optique 1,0 avec les réglages d’acquisition comme indiqué au point 2.1.4. Toutefois, même dans ces conditions, nous avons constaté que l’éclairement continu requis pour l’acquisition de mode stream ne permettait pas pendant une période d’acquisition pendant 2 min. Une illumination plus forte ou une période d’acquisitions dépassant cette limite vous risquez dans non-spécifiques Ca2 + réponses globales et/ou à l’inhibition des processus invasion bactérienne, probablement lié au photovieillissement. Alors que l’imagerie de Ca2 + réponses locales au cours de cinétique plus longue peut être possible avec des moyens de détection plus sensibles, à l’état actuel, que cette imagerie ne peut être effectuée que sur une fraction de temps limité du processus d’invasion de Shigella 15 min. Pour couvrir l’ensemble du processus, nous avons réalisé des séries successives d’acquisitions en continu de 2 min. Cet intervalle de temps est suffisant pour suivre les initiales Ca2 + réponses locales et globales dès le début du processus d’infection et d’établir des différences significatives dans la souche sauvage de Shigella et ses mutants isogéniques ipgD16. Le développement d’une caméra très sensible peut-être permettre des chercheurs pour réduire l’intensité de l’éclairage de fluorophore et exécuter local Ca2 + imagerie sur des périodes prolongées, afin d’améliorer la résolution temporelle de plus Ca transitoire2 + signaux.
Corrélation spatiale entre Ca2 + réponses locales et sites d’invasion bactérienne :
En raison de l’aspect local des événements de signalisation induites par des microorganismes, il est important d’étudier des signaux les de Ca2 + en ce qui concerne leur association avec les sites des interactions hôte-pathogène cellule. Cela a soulevé deux types de considérations. Tout d’abord, toutes les cellules ne peuvent pas être infectés, et tous les micro-organismes ne peuvent pas déclencher signalisation. Pour les Shigella, seule une minorité des bactéries déclencher une invasion, et toutes les cellules ne peuvent pas être infectés. Dans nos expériences, ministère de l’intérieur a été ajusté afin que les lymphocytes forment un 0,7 - 1 objectif de l’invasion bactérienne. Un nombre plus élevé de foyers/cellule peut conduire à une interférence possible pour l’analyse des événements individuels invasion et, à l’inverse, un nombre plus faible peut rendre l’analyse trop complexe, en particulier lorsque l'on étudie les cellules transfectées transitoirement. Nous avons trouvé que l’efficacité de transfection doit atteindre au moins 30 % des cellules pour faire baisser le nombre d’expériences répétées à un nombre gérable. Deuxièmement, Shigella invasion sites peuvent être facilement détectés par microscopie à contraste de phase. Pour d’autres processus, on pourraient servir les autres méthodes de détection par fluorescence. Un aspect important, cependant, est que dans des configurations plus expérimentales, le flux d’acquisition requis pour l’imagerie locale de Ca2 + s’oppose autres types d’acquisition au cours de la période. Cela implique que le processus analysé ne devrait pas montrer mouvement important au cours de ce flux pour permettre leur corrélation spatiale avec les signaux locaux de Ca2 + . Alors que c’est le cas pour les événements d’invasion de Shigella qui localiser sur la même zone de cellules sur plusieurs minutes, processus fortement motiles peuvent ne pas être gérables ou exigeraient probablement plus courts flux d’acquisition.
Notation et l’analyse des réponses du Ca2 + locales :
Alors que Ca2 + réponses globales sont facilement détectés, la détection de Ca2 + réponses locales de faible amplitude et durée nécessite l’optimisation de l’acquisition de signaux fluorescents décrit ci-dessus. Il est également important que les critères stricts sont appliqués afin de différencier les signaux au-dessus des variations d’arrière-plan. En règle générale, nous marquons comme un signal de Ca2 + , soit une augmentation de l’intensité de fluorescence Fluo-4 moyenne qui atteint au moins 3 x les variations de la base de trois acquisitions consécutives de 30 ms. Cette réponse est considérée comme local si une autre région de la cellule montrant la même intensité de fluorescence moyenne ne montre pas une telle augmentation. La notation de Ca2 + réponses locales dans différents échantillons n’entraîne pas de difficultés majeures si ces règles sont systématiquement appliquées. Dans nos mains, la faible fréquence des signaux locaux de Ca2 + associée à l’invasion bactérienne et allant de 5 à 20 % de la cellule analysée représenté un obstacle majeur. Au-delà des variations biologiques détaillées dans le paragraphe précédent, le fait que seulement un flux de données correspondant à une fraction du processus analysés invasion aussi contribue à cette basse fréquence. En raison de cette faible fréquence, établissant des différences entre les échantillons peut-être impliquer effectuant un essai de puissance sur des expériences pilotes pour estimer la taille de l’échantillon pour atteindre une signification statistique.
Applications futures :
Nous croyons que les protocoles fonctionné pour analyser les locaux Ca2 + des signaux pendant une invasion de Shigella utile d’image Ca2 + réponses locales pour tous les processus qui restent dans l’espace occupera au cours de la période d’acquisition. Ca2 + signalisation est polyvalente, Ca2 + signalisation locale peut être plus pertinente pour les processus qui se produisent dans le plasma ou les membranes intracellulaires, où résident les sources de point initiales des signaux. Ainsi, lors des interactions de la cellule hôte microbien, Ca2 + signaux locaux peuvent être pertinentes aux processus adhésifs ou invasifs à la membrane plasmique ou survenant à membranes intracellulaires pathogène contenant des vacuoles. En revanche, les protocoles que nous avons conçu pour analyser les signaux2 + Ca globales sur la cinétique de l’étendue de l’infection peuvent être pertinentes pour les processus qui influent sur la physiologie cellulaire générale lors d’une infection, tels que le règlement d’un programme de transcription ou les voies de la mort cellulaire par des agents pathogènes.
Déclarations de divulgation
Les auteurs n’ont rien à déclarer.
Remerciements
Nous remercions Thomassin Jenny-Lee pour son aide dans l’édition du manuscrit. Le travail a été soutenu par l’ANR accorde MITOPATHO et PATHIMMUN, des subventions du Labex Memolife et Shigaforce de IDEX PSL. Chunhui Sun est récipiendaire d’une bourse de doctorat de la China Scholarship Council. Laurent Combettes et Guy Tran Van Nhieu sont lauréats d’une bourse de WBI-France Tournesol programme N ° 31268YG (Wallonie-Bruxelles International, Fonds de la Recherche Scientifique, Ministère Français des Affaires étrangères et européennes, Ministère de l’enseignement supérieur et de la Recherche dans le cadre des Partenariats Hubert Curien).
matériels
| Name | Company | Catalog Number | Comments |
| Fluo-4 AM | Invitrogen | F14201 | |
| Metamorph version 7.7 | Universal Imaging | ||
| CoolLED illumination system pE-2 | Roper Scientific | ||
| micro-dish 35 mm, high | IBIDI | 81156 | |
| Trypticase Soy (TCS) broth | Thermofisher | B11768 | |
| TCS agar | Thermofisher | B11043 | |
| Congo red | Sigma-Aldrich | 75768 | |
| M90T-AfaE | Sun et al. 2017 | Shigella flexneri serotype V. expressing the AfaE adhesin | |
| ipgD-AfaE | Sun et al. 2017 | isogenic ipgD mutant strain expressing the AfaE adhesin |
Références
- Ashida, H., Ogawa, M., Kim, M., Mimuro, H., Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nature Chemical Biology. 8 (1), 36-45 (2012).
- Berridge, M. J., Lipp, P., Bootman, M. D. The versatility and universality of calcium signaling. Nature Reviews Molecular Cell Biology. 1 (1), 11-21 (2000).
- Strehler, E. E. Plasma membrane calcium ATPases: from generic Ca(2+) sump pumps to versatile systems for fine-tuning cellular Ca(2). Biochemical and Biophysical Research Communications. 460 (1), 26-33 (2015).
- Muallem, S. Decoding Ca2+ signals: a question of timing. Journal of Cell Biology. 170 (2), 173-175 (2005).
- Uhlen, P., Fritz, N. Biochemistry of calcium oscillations. Biochemical and Biophysical Research Communications. 396 (1), 28-32 (2010).
- Carneiro, L. A., et al. Shigella induces mitochondrial dysfunction and cell death in nonmyleoid cells. Cell Host & Microbe. 5 (2), 123-136 (2009).
- Horng, T. Calcium signaling and mitochondrial destabilization in the triggering of the NLRP3 inflammasome. Trends in Immunology. 35 (6), 253-261 (2014).
- Galan, J. E., Lara-Tejero, M., Marlovits, T. C., Wagner, S. Bacterial type III secretion systems: specialized nanomachines for protein delivery into target cells. Annual Review of Microbiology. 68, 415-438 (2014).
- Ashida, H., Mimuro, H., Sasakawa, C. Shigella manipulates host immune responses by delivering effector proteins with specific roles. Frontiers in Immunology. 6, 219 (2015).
- Tran Van Nhieu, G., et al. Actin-based confinement of calcium responses during Shigella invasion. Nature Communications. 4, 1567 (2013).
- Niebuhr, K., et al. Conversion of PtdIns(4,5)P(2) into PtdIns(5)P by the S. flexneri effector IpgD reorganizes host cell morphology. The EMBO Journal. 21 (19), 5069-5078 (2002).
- Konradt, C., et al. The Shigella flexneri type three secretion system effector IpgD inhibits T cell migration by manipulating host phosphoinositide metabolism. Cell Host & Microbe. 9 (4), 263-272 (2011).
- Friedrich, P. The intriguing Ca2+ requirement of calpain activation. Biochemical and Biophysical Research Communications. 323 (4), 1131-1133 (2004).
- Thomas, D., et al. A comparison of fluorescent Ca2+ indicator properties and their use in measuring elementary and global Ca2+ signals. Cell Calcium. 28 (4), 213-223 (2000).
- Sun, C. H., et al. The Shigella type III effector IpgD recodes Ca2+ signals during invasion of epithelial cells. The EMBO Journal. 36 (17), 2567-2580 (2017).
- Allaoui, A., Menard, R., Sansonetti, P. J., Parsot, C. Characterization of the Shigella flexneri IpgD and IpgF genes, which are located in the proximal part of the mxi locus. Infection and Immunity. 61 (5), 1707-1714 (1993).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.