
Method Article
成像 Ca2 +在志贺菌感染上皮细胞期间的响应
摘要
在这里, 我们提出的协议, 可视化钙 (Ca2 +) 的反应引起的 HeLa 细胞感染, 由志贺菌。通过对 ca2 +荧光探针的细菌感染和成像参数进行优化, 提出了由细菌引起的非典型全局和局部 Ca2 +信号在大范围的感染动力学中的特征。
摘要
Ca2 +是所有已知的蜂窝进程中普遍存在的一个离子。尽管全局 Ca2 +响应可能会影响细胞的命运, 但在自由 Ca2 +胞浆浓度的局部变化中, 与内部存储区的释放有关, 或者通过等离子膜通道流入, 调节皮层细胞的过程。附着或侵入宿主细胞的病原体触发了宿主血浆膜肌动蛋白细胞骨架的重组, 这可能会影响全局和局部 Ca2 +信号。由于这些事件可能以伪随机方式出现在扩展动力学上, 因此, 分析由病原体引起的 Ca2 +信号会引发需要解决的重大技术挑战。
在这里, 我们报告的协议, 以检测全球和本地 Ca2 +信号后,志贺菌感染上皮细胞。在这些协议中, 与 Ca2 +荧光探针的激发相关的长时间暴露和光损伤关联的手工制品诊断在志贺菌入侵。在使用化学探针 Fluo-4 对扩展感染动力学过程中的全局胞浆 Ca2 +信号的振幅和频率进行严格分析的过程被实施。
引言
Ca2 +调节所有已知的单元进程, 包括骨架重组、炎症反应和与宿主-病原体相互作用有关的细胞死亡通路1,2,3。在生理条件下, 基底胞浆 Ca2 +浓度低, 在数以百计的 nM 范围内, 但可能会受到瞬变增加后激动剂刺激。这些变化经常表现出振荡行为, 通过泵和渠道在血浆和内质网膜的作用。这些振荡的模式以 ca2 +的周期、持续时间和振幅为特征, 并由单元格进行解密, 然后在被称为 Ca2 +代码4,5 中触发特定响应..在病理条件下, 胞浆 Ca2 +浓度的持续增加可能导致与线粒体膜通透相关的细胞死亡, 并释放亲凋亡或坏死因子6, 7。
志贺菌是细菌性痢疾的致病剂, 通过使用 III. 型分泌系统 (T3SS)8、9, 将效应器注入宿主细胞, 侵入上皮细胞。志贺菌入侵宿主单元格与 T3SS 引发的本地和全局 Ca2 +信号相关联。至于成孔毒素, T3SS translocon 插入宿主细胞膜, 并需要注入 T3SS 效应剂可能负责激活 PLC 和肌醇 (1, 4, 5) trisphosphate (InsP3) 依赖 Ca2 +释放。局部 PLC 刺激和聚合肌动蛋白在志贺菌入侵部位的聚集, 导致超持久的 InsP3-dependent Ca2 +发行10。III. 型效应器 IpgD, 磷脂 45 bisphosphate (PIP2)-4-磷酸酶, 限制了局部 PIP2 量, 从而控制了可编程序控制器生成 InsP3 的可用基板的数量, 这有助于限制本地 Ca2 +细菌入侵站点的响应11,12。这些本地 Ca2 +响应可能会导致在志贺菌入侵站点10的肌动蛋白聚合。但是, 由志贺氏引起的全局 Ca2 +响应是细菌入侵过程不可缺少的, 但会触发连接蛋白 hemichannels 在等离子膜上的打开和胞外 ATP 的释放。车厢。以分泌方式释放的 ATP 将反过来刺激受感染细胞旁边的单元格中的 Ca2 +振荡响应。IpgD 还负责将全局 Ca2 +响应塑造为不稳定的孤立响应, 并具有缓慢的动态。最后, 在长时间的细菌感染后, IpgD 导致 InsP3-mediated Ca2 +信号的抑制。通过对 ca2 +信号的干扰, IpgD 延迟了与 ca2 +相关的钙蛋白酶激活, 导致了焦点黏附结构的拆卸和感染细胞的过早脱离 13.
虽然 Ca2 +信号涉及病机的关键方面, 但微生物的使用引发了一些在经典激动剂研究中没有遇到的技术挑战。此处描述的协议使用常用的荧光 Ca2 +化学指标 Fluo-4, 我们在志贺病毒感染期间设计了用于表征本地 Ca2 +信号的特性。讨论了对这些信号进行检测的关键步骤, 以及为进行定量分析所需的程序, 以确定细菌效应在 Ca2 +信号中的作用。
研究方案
1. 筹备工作
-
细菌制剂
- 板细菌-志贺菌野生型菌株, 表达 AfaE 黏附 (M90T AfaE)-在胰酪胨大豆 (TCS) 琼脂板上含有0.01% 刚果红 (CR) 和孵化他们18小时37摄氏度。
注意: 为了提高其重现性, 在步骤1.1.1 中获得的志贺菌板块储存在4摄氏度, 应在一周内使用, 因为 CR 培养基上的所有菌落最终会随着时间而变红。 - 接种 TCS 肉汤前培养, 从条纹板采摘3红殖民地。
注: 3 个菌落进行接种, 以限制选择单个克隆的可能性, 其毒性质粒经历了基因重组。 - 在16小时在37摄氏度的振动孵化器中培养液体细菌培养 200 rpm。加入氨苄西林, 最后浓度为75毫克/毫升。抗生素浓度不应超过3x 的最小抑制浓度, 因为过量的抗生素可能导致大毒力质粒的损失。
注: 对志贺氏菌毒力质粒的维护应通过电镀铬板上的细菌培养物进行验证, 辅以适当的抗生素浓度。白色菌落的存在表明质粒损失。 - 在1:100 稀释的情况下, 将 TCS 培养与细菌前培养接种。孵化它在37°c 在震动孵化器为2小时在200转每分钟。确保光学密度在 600 nm (OD600nm) 是 0.2-0.4。
- 离心机细菌培养2分钟在 1.3万 x g 在21°c 和并用重悬颗粒在一个等效容量 EM 媒介 (120 毫米氯化钠, 7 毫米氯化钾, 1.8 毫米CaCl2, 0.8 毫米氯化镁2, 5 毫米葡萄糖和25毫米 HEPES, pH 值 = 7.3)。
- 在0.1 的最终 OD600nm中稀释 EM 缓冲器中的细菌悬浮液, 并立即使用, 或将其存储在21摄氏度, 并在下一个60分钟内使用。
- 板细菌-志贺菌野生型菌株, 表达 AfaE 黏附 (M90T AfaE)-在胰酪胨大豆 (TCS) 琼脂板上含有0.01% 刚果红 (CR) 和孵化他们18小时37摄氏度。
-
细胞制剂
- 在 DMEM 中培养 HeLa 细胞, 1 克/升葡萄糖补充10% 胎小牛血清 (FCS), 并将其生长在37摄氏度, 10% CO2。生长 TC-7 生长在 DMEM 与4.5 克/升葡萄糖, 包含 10% FCS 和非必需氨基酸在37°c 孵化器包含 10% CO2。
- 对于细胞的维护, 定期分裂细胞, 以避免与汇合状态相关的生长接触抑制;这对于高 Ca2 +探针加载/转染效率至关重要。
- 在25毫米直径的6井板上, 以 3 x 105细胞的密度为单位, 在 hela 细胞实验前一天, 将 hela 细胞放到无菌的盖玻片上。
- 对于 TC-7 细胞, 胰蛋白酶处理和计数细胞。种子他们在 4 x 105细胞/井和孵化他们 5-7 天在37°c 在 10% CO2-孵化器在实验之前允许极化。
- 每天刷新细胞培养基, 直到实验当天。
注: 玻璃底部的35毫米直径的一次性房间也可以作为替代盖玻片含水井。如果使用后者, 温度控制通过加热的阶段或目标是不足的, 并且一个恒温器孵化室安装在显微镜阶段是必要的。
- 每天刷新细胞培养基, 直到实验当天。
- 实验当天, 取出培养基, 用1毫升 EM 培养基在21摄氏度处冲洗细胞3x。
- 将 3 mM Fluo-4-AM14的单元格加载到 EM 缓冲区中, 其值为21°c 30 分钟。
注: 虽然亚汇合 HeLa 细胞的加载没有引起重大问题, 但极化 TC-7 细胞的加载往往是异构的, 效率低下。对于这些细胞, 我们发现, 增加分散剂-pluronic 酸-在最终浓度为0.1% 和阴离子转运抑制剂 bromosulfophtalein, 增加了加载效率, 最终浓度为20µM 到Fluo-4-AM-containing 他们使用比例化学探针或基因编码的烦恼探头, 要求双采集不符合对本地 Ca2 +响应的映像所需的采集速度。 - 用 em 缓冲器清洗样品 3x, 再在21摄氏度的1毫升 em 缓冲液中孵化, 使 AM 基团水解。立即或在下一个90分钟内使用 Fluo-4 加载的单元格 (保持在21°c)。
2. 感染和图像获取本地 Ca2 +响应
- 将包含 Fluo-4 加载单元格的盖玻片放置在成像室或玻璃底部腔室中。用 EM 缓冲器将成像室或玻璃底部一次性腔内的样品洗净, 去除可能因细胞裂解而产生的化合物。添加1毫升的 EM 缓冲器。
- 将房间放在33摄氏度加热的倒置荧光显微镜阶段。
注意: 此温度被选择为与志贺菌入侵过程兼容的最佳温度与 Ca2 +响应减慢以可视化本地响应之间的折衷。我们使用63X 浸入油目标 (NA = 1.25) 配有相衬环。 - 选择一个显微领域, 并设置采集参数, 包括曝光时间和 binning, 如有必要, 以优化 Fluo-4 荧光信号。
注意: 本地 Ca2 +映像的主要挑战是响应的低振幅和较小的持续时间, 要求至少每30毫秒获取一次. 一些商用的背照明的 EM CCD 或 C MOS 相机提供了所需的使用 Fluo-4 检测本地 Ca2 +响应的灵敏度。检测灵敏度也是一个重要问题, 以限制光损伤或文物, 如自发反应链接到荧光激发的 Fluo-4 在高频率。在这里, LED 照明的470毫微米, 与 480, 40 nm 带通励磁过滤器, 505 nm 分色过滤器, 和 527 @ 30 nm 带通发射, 使用5% 的最大强度与1.0 中性密度过滤器, 以减少这些问题在有限工期的收购下。常规地执行120s 流批处理的本地 Ca2 +信号的分析。在这些低光照条件下, 荧光漂白在2分钟流收购期间没有被观察到。如果使用不同的字段, 则可以执行相同示例的连续收购。一般来说, 在没有细菌刺激的情况下, 第一个采集流是在控制场上进行的, 以确保没有自发反应。如果观察到自发反应, 样品将用 EM 缓冲器冲洗, 直到没有观察到反应。 - 要将细菌添加到样品中, 请从该腔中取出500µL 的 EM 缓冲器, 并添加500µL 在步骤1.1.6 中制备的细菌悬浮液, 以获得最终 OD600nm 0.05, 并适当注意避免移动选定的显微领域;这些体积确保细菌的适当混合和细菌在细胞样品上的均匀分布。
- 对细菌添加进行采集。另外, 在细菌添加后执行10分钟的采集, 这与细菌在使用条件下沉淀到细胞所需的时间相对应。在采集流的末尾, 获取选定字段的相对比图像, 以可视化与细菌侵入点相关的细胞和膜褶边的细菌。
- 按照步骤2.5 中的规定, 重复采集过程, 时间间隔可满足图像获取的持续时间、文件的节省以及选择新字段以覆盖整个过程。
注: 对于志贺菌, 我们发现, 在10分钟细菌沉淀之后, 采集流每5分钟20分钟, 允许覆盖大多数入侵事件。 - 在获取过程的末尾, 添加 ca2 +载体 ionomycin 示例的2µM 的最终浓度, 以确定 ca2 +信号的最大振幅。每 3-5 秒获取图像直到信号稳定, 通常不到10分钟。
- 通过添加 ca2 +离子螯合剂 EGTA 在最终浓度为 10 mM 的情况下, 确定在缺少Ca2 + 的情况下的荧光信号。每 3-5 秒获取图像直到信号稳定, 通常不到10分钟。
注意: 由于全局 Ca2 +变体的动态相对缓慢, 每 3-5 秒 (不在流模式下) 执行这些获取, 在后续处理过程中允许 Ca2 +成像在相同的微观领域。
3. 分析
- 快速胞浆 Ca2 +随时间变化的ΔF/F0的百分比, 其中ΔF 表示感兴趣区域的平均荧光强度 (roi) 和 F0在相同 ROI 中的基线荧光的变化, 其中一个减去单元格的字段的非相关区域。
- 去除不同领域中每个细胞的基底荧光水平, 对应基线水平;由于给定流捕获中本地 Ca2 +响应的低频和相对短的持续时间, 可以明确确定这些级别。
注: 量化与所询问的问题有关的反应的显著特征和研究的致病模型。经典情况下, 考虑了各种参数来分析 Ca2 +信号, 包括显示本地或全局 Ca2 +响应的单元格的频率, 以及这些响应的持续时间、振幅和频率。在志贺菌和局部反应的情况下, 分析的另一个重要方面是与细菌入侵部位的反应的空间关联。
- 去除不同领域中每个细胞的基底荧光水平, 对应基线水平;由于给定流捕获中本地 Ca2 +响应的低频和相对短的持续时间, 可以明确确定这些级别。
- 为了量化显示全球或局部反应的应答细胞的百分比, 在细胞中绘制 ROIs, 时间序列对应 Fluo-4 强度的变化。
- 设置严格的参数以标识本地 Ca2 +响应, 例如, 在平均荧光强度单元基线的背景变化的三倍以上的振幅, 或至少200毫秒的持续时间, 以避免得分为假阳性。
注意: 作为一般规则, 即使小的本地 Ca2 +响应也可以与其配置文件中的任何背景噪音区分开来。ROIs 应该足够大, 能够集成足够的荧光信号, 以便检测局部变化。根据 Fluo-4 检测的强度, 这些 ROIs 可能对应于直径从 2-5 毫米不等的圆。 - 测量同一单元格不同区域中2个区域中平均 Fluo-4 荧光强度的变化, 以确定响应是局部的还是全局的。
注: 通过使用成像软件, 允许在线屏幕显示所选区域的平均荧光强度变化, 从而促进了对大量细胞的分析。商业上可用的成像软件有这样一个选项, 但也可以使用ROI 强度演化插件在冰冷的软件中执行此操作。 - 确定显示本地响应的单元格的百分比。
注意: 这个百分比是从显微镜领域分析的细胞总数计算出来的, 应该有足够的数字来支持使用非参数测试的统计学意义。 - 确定本地响应的持续时间。
注意: 本地 Ca2 +响应可根据其持续时间范围 (即小于500毫秒, 介于5和十年代之间, 或在十年代以上) 进一步区分, 并与志贺氏菌入侵站点关联。
- 设置严格的参数以标识本地 Ca2 +响应, 例如, 在平均荧光强度单元基线的背景变化的三倍以上的振幅, 或至少200毫秒的持续时间, 以避免得分为假阳性。
- 量化响应的振幅。首先, 校准最大 Ca2 +和确定为步骤2.1.7 的单元格背景级别。由于每个单元格的 Fluo-4 负载可能会有所不同, 请对该字段中的单个单元格执行这些决定。
- 将响应的振幅表示为最大响应的相对百分比。
- 使用正常测试进行统计测试, 然后进行参数化测试, 分析样本间响应幅度的潜在差异。
- 根据相应的相衬图像中的细菌诱导膜褶边, 确定这些反应相对于细菌侵入点的相关性。
结果
志贺菌入侵与非典型的持久性本地 Ca2 +响应关联:
根据上述协议, Fluo-4-loaded HeLa 细胞受到了志贺菌的挑战, 并进行了流采集以分析 Ca2 +信号。一个具有代表性的实验显示在图 1中, 其中的时间推移图像系列的荧光强度的 Fluo-4 探针平均在一个区域感兴趣的一个单元, 和相应的相位对比度图像 (图 1A, 左面板)。志贺氏入侵站点的特征是在相对比图像中检测到膜褶边 (图 1A、箭头)。游离胞浆 Ca2 +的非典型局部增加在具有不同振幅和持续时间范围 (从 2.5-5 s (图 1A和1B、箭头) 的志贺氏入侵站点观察, 其次是受感染的全球增加单元格 (图 1B、箭头)。
III 型效应器 IpgD 调节从本地到全局 Ca2 +在 HeLa 细胞中由志贺菌引起的反应:
分析了野生型志贺菌诱发的 Ca2 +信号, 以及 III. 型效应 IpgD (磷脂4、5 bisphosphate 磷酸酶) 的等基因系突变株缺陷, 表明后一种菌株引起的全球和更少不典型的局部反应与长的持续时间 (RATPs) 与 11.3 "4.4 (sem)% 和 40.3 * 7.5 (sem) 对响应全球反应的细胞%% 在 ipgD 突变体的情况下, 分别 (图 1D)15。这个 ipgD 突变体以类似的频率得到局部响应, 如应变。
志贺菌抑制 InsP3-dependent 的全球 Ca2 +在长时间感染动力学过程中的 HeLa 细胞增加:
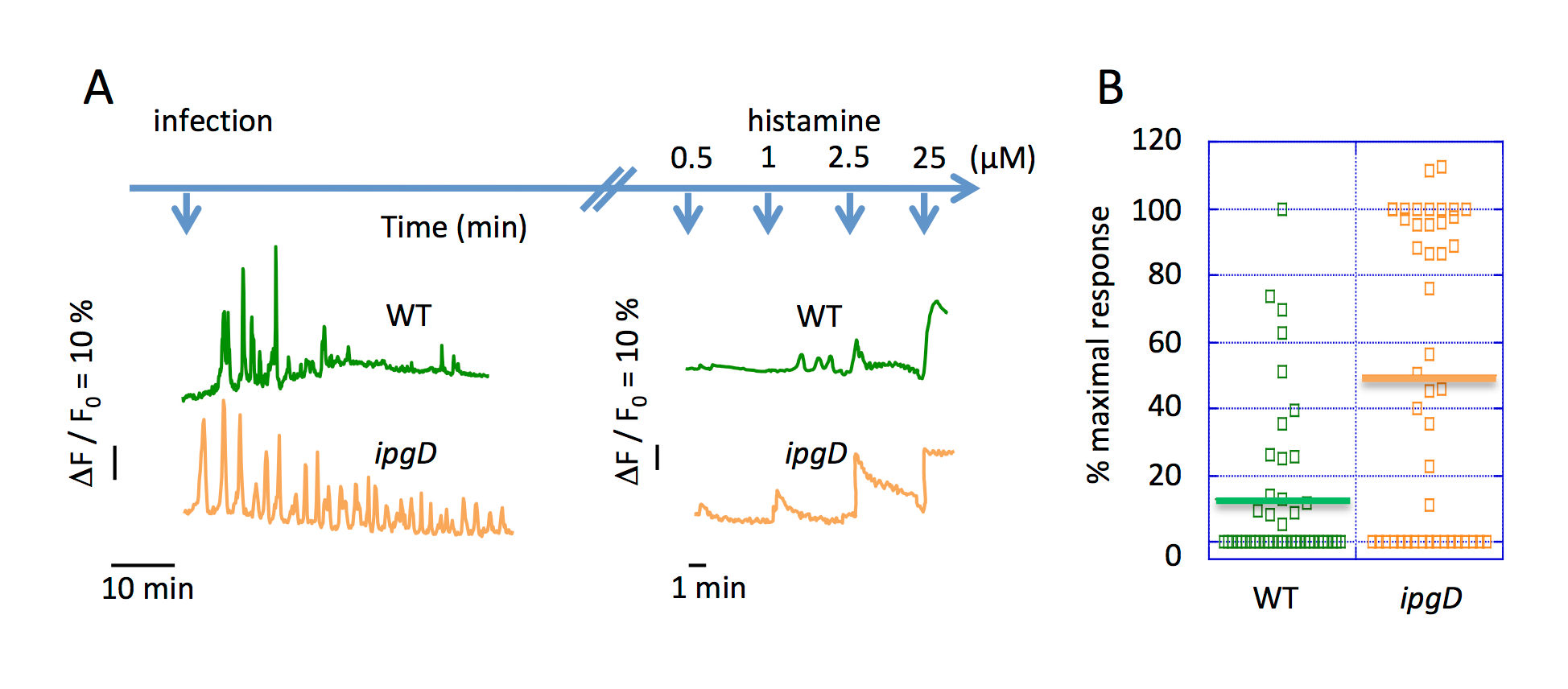
对扩展感染动力学的全局 Ca2 +响应的映像表明, 在使用野类型志贺菌(图 2A) 挑战后, 响应的频率减少了30分钟。与野生型志贺菌15一起感染30分钟的 TC-7 细胞 Ca2+ 反应频率也有类似的减少。对 Ca2 +激动剂组胺的剂量依赖性细胞反应的量化说明了在志贺菌感染晚期的 InsP3-dependent Ca2 +释放的抑制。在感染前30分钟和进一步孵化之后,志贺菌导致非典型隔离 ca2 +响应, 观察到 ca2 +响应的振幅和频率急剧下降 (图 2B)。

图 1.本地和全局 Ca2 +在志贺菌入侵期间的响应.HeLa 细胞加载与 Fluo-4-AM 和挑战的志贺菌。(A) 左面板显示相对比图像。右面板显示了 Fluo-4 荧光平均强度的时间序列, 并在右侧描述了颜色代码。从承购开始的时间在秒, 10 分钟被表明在细菌挑战以后。箭头指示入侵焦点。所描述的圆圈对应于 (B) 中分析的区域, 其中具有相应颜色的跟踪表示基线上 Fluo-4 平均荧光强度的变化。箭头指示本地响应。箭头表示全局响应。(C) 这是一个描述 Ca2 +响应持续时间的方案。水平虚线表示基 Ca2 +级别 (F0);垂直条表示背景变化。垂直虚线表示响应的最大振幅;水平条表示响应的持续时间 (以半最大振幅确定)。(D) 这是响应细胞的百分比, 扫描电镜显示本地响应和全局 Ca2 +响应诱发5分钟后感染的 ipgD志贺菌(深灰色条) 或突变体 (浅灰色条)。RATPs 是本地 Ca2+ 响应, 持续超过5秒. N = 4;> 每一次60细胞。魏氏测试, *P < 0.05;**P < 0.01。请单击此处查看此图的较大版本.
{kind=link}

图 2.抑制全球 Ca2 +的反应在延长的动力学的志贺菌感染。() 上蓝色箭头表示细菌和组胺挑战在指定浓度下的时间刻度.此面板显示了在病毒感染后的单个单元格全局 Ca2 +变体的代表性痕迹, 这些变化发生在由 ipgD (绿) 和突变株 (橙色) 引起的. (B) 此面板显示了相对于最大响应的 Ca2 +响应振幅的百分比, 在0.5 微米组胺的刺激下, 被指定菌株感染的细胞为90分钟。实心水平条表示平均最大百分比。魏氏测试, **P < 0.01。请单击此处查看此图的较大版本.
{kind=link}
讨论
本手稿描述了在志贺菌入侵的相对较短的动力学过程中, 我们设计为遵循本地 ca2 +信号的协议, 以及志贺菌扩展动力学过程中的全局 ca2 +响应。下面, 需要解决的关键问题是优化 Ca2 +信号的检测, 同时尽量减少对生物过程的任何干扰。
化学vs基因编码的 Ca2 +探头:
为了图像本地 Ca2 +的变化, 我们使用了 Fluo-4 化学探针, 因为它的高量子产量和它的快速反应动力学。由于无法执行双波长采集, 因此采集所需的速度也排除了比例探头的使用。然而, 致病微生物的使用可能会干扰使用 Ca2 +化学探针的标准程序。例如, 由志贺氏菌引起的长时间细胞感染可防止任何细胞加载化学探针, 想必是由于病原体引起的宿主细胞血浆膜的改变。由于该探针的细胞质损失, 在长时间感染动力学过程中观察到细胞相关 Fluo-4 荧光的减少。因此, 实现一个控件, 如添加 ionomycin 以确定在收购结束时最大 Ca2 +浓度的荧光, 对于识别响应单元至关重要。此外, 与病原体感染相关的极化肠道上皮细胞似乎难以被载入 Ca2 +探针, 并且需要特定的加载过程。虽然使用基因编码的 Ca2 +记者 (GECR) 可能有助于解决其中的一些问题, 但也会引入其他挑战。宿主细胞系统的转染效率可能是一个严重的限制, 特别是如果它叠加的感染率很低。此外, 对大多数 GECRs 的 ca2 +变体的响应特性与本地 Ca2 +分析所需的快速动力学不兼容。最后, GECR 的表达可能会干扰病原体介导的过程。我们不会讨论 GECR 在宿主-病原体相互作用期间对 Ca2 +信号进行研究的使用。最近和正在进行的各种 "快速反应" GECR 的工程可能值得研究人员在未来的研究中重新使用。
在细菌感染期间对本地 Ca2 +响应的定时获取:
局部 Ca2 +响应的成像需要高速的图像采集, 这会使 Fluo-4 探针持续的荧光激发。由于与高频采集有关的光损伤, 荧光励磁光的强度保持在达到足够的信噪比且曝光时间不超过30毫秒的情况下是至关重要的. 我们有良好的结果使用 LED 系统的最大强度的 5%, 结合了1.0 光密度滤波器与采集设置, 如步骤2.1.4 所述。然而, 即使在这些条件下, 我们发现, 流模式获取所需的连续光照不允许超过2分钟的购置期。更强的光照或获取周期超过此限制可能导致非特定的全局 Ca2 +响应和/或抑制细菌入侵过程, 大概与光损伤。虽然对较长的动力学的局部 Ca2 +响应的成像可能具有更灵敏的检测手段, 但目前状态下这种成像只能在15分钟的志贺菌入侵过程的有限时间段内执行。为了涵盖整个过程, 我们连续进行了一系列2分钟的流式收购。此时间间隔足以在感染过程开始时遵循初始本地和全局 Ca2 +响应, 并在志贺菌野型菌株及其等基因系 ipgD突变16 中建立显著差异。高度敏感的照相机的开发可能允许研究人员进一步降低荧光照明的强度, 并在较长的时间内执行本地 Ca2 +映像, 从而提高更多瞬态 ca2 + 的时间分辨率信号。
本地 Ca2 +响应与细菌入侵站点之间的空间相关性:
由于微生物引起的信号事件的局部方面, 研究本地 Ca2 +信号与病原体-宿主细胞相互作用的地点有关, 是很重要的。这提出了两种考虑因素。首先, 所有细胞可能不会被感染, 所有的微生物可能不会触发信号。对于志贺菌, 只有少数细菌触发入侵, 所有细胞都可能不会被感染。在我们的实验中, 语言的调整, 使一个细胞形成 0.7-1 的焦点细菌入侵。更多的病灶/细胞可能导致对个体入侵事件分析的可能干扰, 反之, 较低的数字可能会使分析过于复杂, 特别是在研究瞬时转染细胞时。我们发现, 转染效率需要达到至少30% 的细胞, 以降低复制实验的数量, 以可管理的数字。其次, 通过相衬显微镜可以很容易地检测到志贺菌入侵部位。对于其他过程, 可以使用其他基于荧光的检测方法。但是, 一个重要的方面是, 在大多数实验性的设置中, 本地 Ca2 +映像所需的捕获流排除了期间内其他类型的获取。这意味着分析的过程不应在流期间显示显著的运动, 以允许它们与本地 Ca2 +信号的空间相关性。虽然在几分钟内本地化到同一单元格区域的志贺氏入侵事件是如此, 但高度活动的过程可能无法管理, 或者可能需要较短的捕获流。
对本地 Ca2 +响应的评分和分析:
虽然可以很容易地检测到全局 ca2 +响应, 但对于小幅度和持续时间的局部 ca2 +响应的检测需要优化上面描述的荧光信号的获取。同样重要的是, 采用严格的标准来区分背景变化之上的信号。作为一条规则, 我们将分数作为 Ca2 +信号增加平均 Fluo-4 荧光强度, 达到至少3x 三连续 30 ms 收购的基线变化。如果显示相同平均荧光强度的单元格的另一个区域没有显示这种增加, 则该响应被认为是局部的。如果系统地应用这些规则, 则在各种示例中对本地 Ca2 +响应的评分不应造成重大困难。在我们的手中, 低频率的本地 Ca2 +信号相关的细菌入侵和范围从 5-20% 的细胞分析代表了一个主要障碍。除了前一段所详述的生物变异之外, 仅一条与分析的入侵过程的一小部分相对应的流也会导致这种低频现象。由于这种低频, 建立样本之间的差异可能牵连到进行试验试验的功率测试, 以估计样本大小, 以达到统计学意义。
未来应用:
我们认为, 在志贺菌入侵期间, 分析本地 ca2 +信号的协议将有助于图像本地 ca2 +对在获取期间内保持空间限制的所有进程的响应。尽管 Ca2 +信号是通用的, 但本地 Ca2 +信号可能与在初始点信号源所在的等离子或胞内膜中发生的进程最相关。因此, 在微生物宿主细胞相互作用的过程中, 局部 Ca2 +信号可能与等离子膜上的粘附或侵入过程有关, 或发生在含有病菌的液泡的胞内膜中。另一方面, 我们设计用于分析全局 Ca2 +信号超过扩展感染动力学的协议可能与在感染期间影响一般细胞生理学的过程相关, 例如对转录程序的调节。或细胞死亡通路的病原体。
披露声明
作者没有什么要申报的。
致谢
我们感谢珍妮-李. Thomassin 在编辑手稿方面的帮助。这项工作得到了情报局赠款 MITOPATHO 和 PATHIMMUN 的支持, Labex Memolife 和人民党 IDEX Shigaforce 提供赠款。春晖孙是来自中国奖学金委员会的博士助学金的接受者。劳伦 Combettes 和家伙搬运车 Nhieu 是 WBI-法国交换 Tournesol 计划 N°31268YG (Wallonie 布鲁塞尔国际, 全宗研究所科学研究, 部法国, étrangères et européennes, 部 de 的接受者。l ' Enseignement 高级研究所 dans 乐干部 des Partenariats 休伯特克瑞恩)。
材料
| Name | Company | Catalog Number | Comments |
| Fluo-4 AM | Invitrogen | F14201 | |
| Metamorph version 7.7 | Universal Imaging | ||
| CoolLED illumination system pE-2 | Roper Scientific | ||
| micro-dish 35 mm, high | IBIDI | 81156 | |
| Trypticase Soy (TCS) broth | Thermofisher | B11768 | |
| TCS agar | Thermofisher | B11043 | |
| Congo red | Sigma-Aldrich | 75768 | |
| M90T-AfaE | Sun et al. 2017 | Shigella flexneri serotype V. expressing the AfaE adhesin | |
| ipgD-AfaE | Sun et al. 2017 | isogenic ipgD mutant strain expressing the AfaE adhesin |
参考文献
- Ashida, H., Ogawa, M., Kim, M., Mimuro, H., Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nature Chemical Biology. 8 (1), 36-45 (2012).
- Berridge, M. J., Lipp, P., Bootman, M. D. The versatility and universality of calcium signaling. Nature Reviews Molecular Cell Biology. 1 (1), 11-21 (2000).
- Strehler, E. E. Plasma membrane calcium ATPases: from generic Ca(2+) sump pumps to versatile systems for fine-tuning cellular Ca(2). Biochemical and Biophysical Research Communications. 460 (1), 26-33 (2015).
- Muallem, S. Decoding Ca2+ signals: a question of timing. Journal of Cell Biology. 170 (2), 173-175 (2005).
- Uhlen, P., Fritz, N. Biochemistry of calcium oscillations. Biochemical and Biophysical Research Communications. 396 (1), 28-32 (2010).
- Carneiro, L. A., et al. Shigella induces mitochondrial dysfunction and cell death in nonmyleoid cells. Cell Host & Microbe. 5 (2), 123-136 (2009).
- Horng, T. Calcium signaling and mitochondrial destabilization in the triggering of the NLRP3 inflammasome. Trends in Immunology. 35 (6), 253-261 (2014).
- Galan, J. E., Lara-Tejero, M., Marlovits, T. C., Wagner, S. Bacterial type III secretion systems: specialized nanomachines for protein delivery into target cells. Annual Review of Microbiology. 68, 415-438 (2014).
- Ashida, H., Mimuro, H., Sasakawa, C. Shigella manipulates host immune responses by delivering effector proteins with specific roles. Frontiers in Immunology. 6, 219(2015).
- Tran Van Nhieu, G., et al. Actin-based confinement of calcium responses during Shigella invasion. Nature Communications. 4, 1567(2013).
- Niebuhr, K., et al. Conversion of PtdIns(4,5)P(2) into PtdIns(5)P by the S. flexneri effector IpgD reorganizes host cell morphology. The EMBO Journal. 21 (19), 5069-5078 (2002).
- Konradt, C., et al. The Shigella flexneri type three secretion system effector IpgD inhibits T cell migration by manipulating host phosphoinositide metabolism. Cell Host & Microbe. 9 (4), 263-272 (2011).
- Friedrich, P. The intriguing Ca2+ requirement of calpain activation. Biochemical and Biophysical Research Communications. 323 (4), 1131-1133 (2004).
- Thomas, D., et al. A comparison of fluorescent Ca2+ indicator properties and their use in measuring elementary and global Ca2+ signals. Cell Calcium. 28 (4), 213-223 (2000).
- Sun, C. H., et al. The Shigella type III effector IpgD recodes Ca2+ signals during invasion of epithelial cells. The EMBO Journal. 36 (17), 2567-2580 (2017).
- Allaoui, A., Menard, R., Sansonetti, P. J., Parsot, C. Characterization of the Shigella flexneri IpgD and IpgF genes, which are located in the proximal part of the mxi locus. Infection and Immunity. 61 (5), 1707-1714 (1993).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。