
Method Article
Um ensaio comportamental para investigar o papel da memória espacial durante uma defesa instintiva em ratos
Neste Artigo
Resumo
Este protocolo descreve um ensaio comportamental, com base no teste de labirinto de Barnes, para estudar como instintiva defensiva ações são modificadas pelo conhecimento do ambiente espacial.
Resumo
Evolução selecionou um repertório de comportamentos defensivos que são essenciais para a sobrevivência em todas as espécies animais. Esses comportamentos são muitas vezes ações estereotipadas que suscitou em resposta a estímulos sensoriais intrinsecamente contrário, mas seu sucesso requer flexibilidade suficiente para adaptar a diferentes ambientes espaciais, que podem mudar rapidamente. Aqui, descrevemos um ensaio comportamental para avaliar a influência do conhecimento espacial em comportamentos defensivos em camundongos. Adaptámos o ensaio de memória espacial de labirinto Barnes amplamente usado para investigar como os ratos navegam para um abrigo durante respostas de fuga aos estímulos sensoriais intrinsecamente contrário em um ambiente de romance, e como eles se adaptam às mudanças agudas no ambiente. Este novo ensaio é uma paradigma Etológica que não requer treinamento e explora os padrões de exploração natural e estratégias de navegação em camundongos. Propomos que o conjunto de protocolos descritos aqui são um meio poderoso de estudo dos comportamentos objetivo direcionado e acionadas por estímulo de navegação, que deve ser de interesse para ambos os campos de comportamentos instintivos e memória espacial.
Introdução
Comportamentos defensivos instintivos são amplamente considerados hardwired-respostas aos estímulos, tais como o movimento de C-início em peixes e anfíbios, que move o animal longe de uma fonte de ameaça1. No entanto, comportamentos defensivos podem ser mais adaptáveis se flexìvel levam em conta as informações sobre o ambiente atual. Um exemplo de tal flexibilidade é o interruptor de fuga para congelamento mostrado por roedores quando confrontado com a ameaça, dependendo do conhecimento prévio sobre a presença ou ausência de um abrigo no ambiente2,3. Outros exemplos de flexibilidade em comportamentos inatas incluem adapta o limiar de início de voo ou escapar de velocidade dependendo da distância entre uma presa e seu abrigo4, distância anteriores e a ameaça de5,6, experiência 11 , 12, bem como selecionando diferentes estratégias defensivas, dependendo das propriedades sensoriais do estímulo aversive7, ou mesmo suprimir o comportamento defensivo face a concorrentes motivações como fome8, 9,10. A dependência de seleção de ação no conhecimento adquirido sobre as características espaciais do ambiente faz instintivos comportamentos defensivos em camundongos um poderoso modelo para o estudo objetivo seleção, memória espacial e navegação. Aqui, descrevemos a adaptação de uma tarefa comportamental comumente usada, a memória espacial de labirinto (BM) de Barnes do ensaio13, para determinar a influência do ambiente espacial na seleção de ação defensiva em ratos e suas estratégias de navegação quando fugindo em direção um abrigo.
O labirinto de Barnes padrão usado para estudar a memória espacial e aprendizagem em ratos consiste em uma plataforma de diâmetro circular ~ 90 cm com 20 furos igualmente espaçados, dos quais 19 estão fechadas, e um leva a um abrigo subterrâneo que ratos procuram a fim de evitar o campo aberto ambiente da plataforma. Estímulos aversive fracos (campainha, luz brilhante, ventilador) são frequentemente usados continuamente durante todo o ensaio para tornar o ambiente contrário e assim promover a entrada para o abrigo de14. No ensaio mais comumente usados15,16, o animal tem um julgamento de habituação onde pauta-se para o abrigo manualmente pelo experimentador imediatamente após ser posicionado na plataforma. Isto é seguido por um período de aquisição de 4 dias, onde cada dia que o mouse é permitido navegar livremente no labirinto por 3 min, após o qual novamente manualmente pauta-se para o abrigo se não alcançar, durante o período de exploração. A última fase do ensaio é uma sonda de julgamento sobre o 5º dia (embora uma memória a longo prazo de sete dias julgamento é também comumente realizada) quando o animal explora o labirinto com todos os buracos fechados. Aprendizagem e a longo prazo de memória são quantificados pelo tempo gasto para encontrar o abrigo e pokes nos buracos errados durante o período de aquisição e quando passei perto do furo de abrigo fechado no julgamento de sonda. Resultados típicos mostram uma diminuição do número de erros e latência para alcançar o abrigo durante a aquisição e uma proporção acima-oportunidade de tempo gasto no quadrante contendo o buraco fechado alvo da sonda experimental15.
Enquanto diversas variantes do ensaio BM têm sido descritas anteriormente17,18,19, o paradigma que descrevemos aqui tem três mudanças fundamentais de ensaio padrão. Primeiro, o animal é deixado para explorar o labirinto e encontrar o abrigo por conta própria, e teste é feito na mesma sessão, logo após o abrigo foi encontrado, durante os períodos onde o animal está envolvido em comportamento exploratório. Embora essa configuração não testar a memória de longo prazo, do local de abrigo, ele é projetado para ser um cenário naturalista que imita a exploração do novo território, sob a ameaça de predação. Além disso, permite testes de como animais se adaptar às mudanças agudas no ambiente, tais como mudanças bruscas na paisagem. Em segundo lugar, um aspecto fundamental do nosso ensaio é que o experimentador nunca força o animal dentro ou fora do abrigo, que pode desorientar o animal e excluir caminho integração20 como uma estratégia de navegação viável21. Integração do caminho é uma estratégia de navegação que utiliza sinais de auto-movimento, como pistas proprioceptivas e vestibulares, decorrentes da integração da saída do motor, para atualizar a posição atual do animal e navegar para um objetivo, que não é possível se o animal é deslocado passivamente pelo experimentador. Em terceiro lugar, usamos intrinsecamente contrário visual22 e estímulos auditivos23 para eliciar a fuga para o abrigo, que é facilmente distinguido do comportamento de forrageamento em curso e permite a avaliação e quantificação das estratégias específicas de navegação usado durante a defesa de ameaças iminentes. Propomos que este ensaio será útil para dissecar o papel da memória espacial na seleção e implementação de comportamentos defensivos, e mais geralmente para o mais amplo estudo de navegação objetivo-dirigido e a curto prazo memória espacial. Os protocolos descritos aqui foram introduzidos por Vale et al. em 2017, a que nos referimos aos leitores para mais na discussão de profundidade sobre a justificativa para as experiências e resultados.
Protocolo
Todos os experimentos foram realizados sob o ato de UK animais (procedimentos científicos) de 1986 (70/7652 PPL) após a aprovação ética local.
1. criação de aparato comportamental
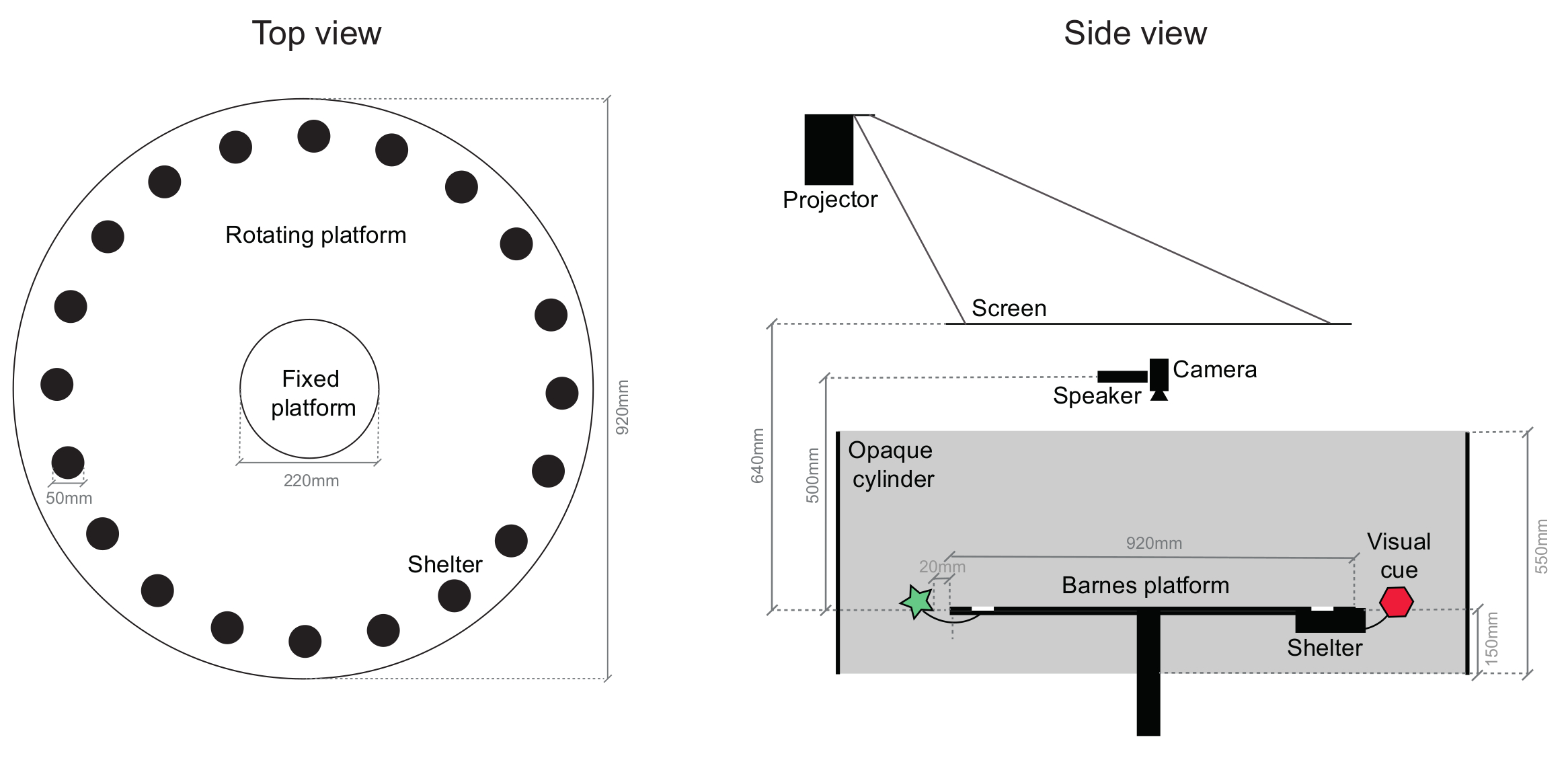
Nota: O principal componente do aparato comportamental é uma variante da Barnes labirinto13 consiste em uma plataforma circular acrílica (92 cm de diâmetro) branca com 20 equidistantes, 5 cm de diâmetro, orifícios circulares, localizado radialmente, 5 cm da borda da plataforma . 19 delas são para ser fechada com tampa de plástico preta (5 mm de profundidade), enquanto o restante buraco leva a um abrigo de Perspex preto (dimensões 15 x 5,8 x 4,7 cm) que os ratos podem facilmente entrar e sair. A área central da arena consiste em uma plataforma circular fixa (22 cm de diâmetro) e a periferia é montada sobre um quadro que permite mais de 360 graus de rotação através de um motor de passo controlado por computador. A plataforma é elevada a 45 cm do chão para evitar que os ratos descer. Veja o desenho esquemático na Figura 1.
- Anexar sugestões de visual para a periferia da plataforma (2D ou 3D símbolos com diferentes formas, cores e padrões), pelo menos a 2 cm da borda para evitar ratos de alcançá-los.
Nota: Exemplos de indicações visuais são simples quadrados azuis de 262 cm2ou círculos (diâmetro de 15,5 cm) cheios de listras diagonais de roxas. - Adicione ~ 2 g do fundamento da gaiola do rato de casa no interior do abrigo para servir como uma sugestão de local olfativa. Opcionalmente, posicione uma luz emitida por diodo (LED) na entrada do abrigo para fornecer uma indicação de marco visual local.
- Para isolar o mouse externo, indicações visuais distais, montar uma parede cilíndrica ou octogonal, estendendo-se 40 cm acima e 15 cm abaixo da superfície da plataforma, em torno da arena, posicionada a 10 cm de distância da borda da plataforma.
- Posicione um altofalante de ultra-som 50 cm acima do centro da plataforma, para fornecer estímulos auditivos aversive.
- Coloque uma tela translúcida 64 cm acima da arena para trás-projeto sobrecarga aversive estímulos visuais usando um projetor.
- O fornecido pelo projector acenderá o aparato comportamental; calibrá-lo para que haja 5-10 lx no centro da superfície da plataforma. Coloque luzes infra-vermelhas e uma câmera infravermelha acima da plataforma, com um filtro de passa-tempo (> 700 nm) para evitar tremulação de projetor no vídeo adquirido.
- Controle a posição do mouse on-line, usando um algoritmo apropriado ou software disponível comercialmente. Para controlar os ratos pretos em uma arena de branca, uma operação simples de limiarização seguida pelo cálculo do centroide da máscara é apropriada.
- Idealmente, coloque o aparelho comportamental dentro de uma caixa de som - e à prova de luz, para evitar que sinais externos que afetam a experiência.
2. fornecimento de estímulos intrinsecamente contrário
Nota: Os estímulos descritos abaixo podem ser gerados com uma vasta gama de pacotes de software diferentes, incluindo o LabVIEW e Matlab.
- Estímulo visual
- Projeto dos estímulos para a tela posicionado 64 cm acima da arena. O estímulo é composto por um círculo escuro linearmente em expansão em um fundo cinza22 (contraste Weber =-0.98, luminância = 7,95 cd/m2).
Nota: O círculo padrão idealmente deve ser centralizado diretamente acima da posição do mouse, subtending num ângulo visual de 2,6 ° pelo aparecimento e expande-se linearmente em 224 ° /s mais de 200 ms para 47,4 °, no qual permanece por 250 ms antes de deslocamento. - Quando entregar o estímulo visual, centralizá-la acima do mouse. Isto pode ser conseguido esperando até que o mouse passa por uma região previamente definida na arena, ou rastreamento da posição do mouse on-line e usando as coordenadas de localização para definir o centro do círculo.
Nota: Embora sugerimos projetando o estímulo visual diretamente acima do rato, comportamentos defensivos podem também ser provocados se projeta-se o mesmo estímulo à frente o mouse (enquanto ainda paralela ao chão). Em nossa experiência, este é o caso, pelo menos até 30 cm à frente da posição do mouse.
- Projeto dos estímulos para a tela posicionado 64 cm acima da arena. O estímulo é composto por um círculo escuro linearmente em expansão em um fundo cinza22 (contraste Weber =-0.98, luminância = 7,95 cd/m2).
- Estímulo auditivo
- Desencadear um estímulo que consiste de um comboio de três frequência modulada acima-varreduras de 17 a 20 kHz23 mais de 3 s, durando 9 s no total. O nível de pressão sonora (SPL), medido no nível da plataforma, deve ser em um intervalo entre 73 e 78 dB.
- Ligar uma placa de som e um amplificador ao altifalante ultra-som posicionado acima da arena para entregar o estímulo auditivo.
3. animais:
- Use Camundongos C57BL/6J masculinos ou femininos, entre 6 e 24 semanas de idade. Manter que os animais em uma luz h 12 ciclo com livre acesso a comida e água.
- Certifique-se de que os ratos são único abrigado pelo menos 72 h antes do ensaio comportamental e teste-os durante a fase de luz da luz do ciclo.
Nota: Nós achamos que o comportamento exploratório de base dos ratos alojados em single é mais comparável em animais do que em ratos alojados em grupo. Além disso, animais com implantes cirúrgicos para gravação de atividade neural ou manipulação frequentemente precisam ser único abrigado, e, portanto, pode ser benéfico ter conjuntos de dados do controle nas mesmas condições de habitação.
4. padrão ensaio comportamental:
- Meticulosamente, limpe a superfície da plataforma com 70% de etanol ou ácido acético para remover indesejadas sugestões olfactory. Lave o abrigo com água e lave-o com etanol a 70%, após o qual ele deve ser lavado com água novamente para diminuir o odor de etanol.
- Seque bem tanto a plataforma e o abrigo, como o mouse pode evitar entrar no abrigo se está molhado.
- Randomize a localização do abrigo na plataforma para cada julgamento, rodando a plataforma antes do experimento. O local de abrigo será ancorado na arena, mas será rotacionado em relação ao compartimento do aparato comportamental.
- Trazer o rato para a sala de experiências em sua casa-gaiola e coloque a gaiola em cima da plataforma do teste por um período de aclimatação de 10 min.
- Remover o rato de sua casa-gaiola e evitar recuperando pela cauda: Copa-lo ou deixá-lo subir um item de enriquecimento que pode ser levantado. Pegar ratos pela cauda foi mostrar para aumentar os níveis de ansiedade e assim pode afetar as respostas comportamentais para ameaça24.
- Delicadamente, coloque o mouse no centro da arena e começar a aquisição de vídeo.
- Permita, por um período de habituação de 7 min, durante o qual nenhum estímulo contrário é apresentado. Certifique-se de que o mouse tem tempo suficiente para encontrar e entrar no abrigo pelo menos uma vez. Considerá-la como uma entrada se todos os quatro membros estão dentro do abrigo. 7 min deve ser suficiente para a maioria dos ratos encontrar e entrar no abrigo, mas caso isso não aconteça, prorrogar o período de habituação por períodos adicionais de 5 min até que tenha verificada pelo menos uma entrada.
- Acionar a entrega do estímulo contrário (auditivo ou visual) ou manualmente, observando o feed de vídeo ao vivo, ou automaticamente pelo rastreamento da posição do animal on-line e pré-definir uma região de interesse na arena para desencadear o estímulo-entrega uma vez que o animal chega-lo. O estímulo visual é normalmente entregue diretamente acima do animal usando o vídeo on-line rastreamento coordenadas.
- Para obter mais do que uma reação de fuga na mesma sessão, deixe o mouse abandonar voluntariamente o abrigo. Se o mouse não escapou com êxito para o abrigo após o último estímulo contrário, espere pelo menos 60 s antes de apresentar outro estímulo.
- Depois que terminar o ensaio, devolver o animal para sua casa-gaiola. No caso, são necessários mais ensaios do mouse mesmo, espere pelo menos 48 h antes de refazer o teste.
- Limpe a plataforma e o abrigo, conforme descrito na etapa 4.1 e 4.2 antes de testar o próximo animal.
Nota: As etapas 5, 6 e 7 são variações independentes do ensaio padrão comportamental e cada um pode ser realizado individualmente.
5. «estratégia de navegação» ensaio comportamental:
Nota: O objetivo deste teste é determinar que cues mouses usam para orientar o comportamento de fuga. Neste ensaio, a parte externa móvel da plataforma é girada, enquanto o mouse está no centro da arena fixa, que desloca radialmente o abrigo, as sugestões olfactory, localizadas no interior do abrigo e indicações visuais proximais anexadas à plataforma. Se o mouse está seguindo qualquer uma das dicas que têm sido giradas para navegar para o abrigo, ele escapará corretamente para o novo local do abrigo, mas se ele usa quaisquer outras sugestões que permaneceram no lugar, navegará até o local de abrigo anterior.
- Execute as etapas 4.1 a 4.8 do padrão ensaio comportamental descrito acima, provocando pelo menos uma resposta de voo para ser usado como linha de base.
- Deixe o mouse ativamente sair do abrigo e esperar por ele para navegar até a parte central da plataforma. Coloque um pequeno prato com fundamento no centro para incentivar a exploração e manter o animal na plataforma central suficientemente longo para permitir a rotação da parte externa da plataforma. Adicione o prato e roupa de cama enquanto o mouse está dentro do abrigo e após a fuga bem sucedida, que precede a rotação da plataforma. Se o rato não entrar o abrigo durante a resposta de fuga, espere por ele para entrar voluntariamente o abrigo e em seguida, adicione o prato e roupa de cama.
- Use o motor de passo para girar a plataforma pelo menos 36 graus (2 furos) e imediatamente use um estímulo sensorial para eliciar uma resposta de fuga. É possível que a rotação aciona um alarme e nem inicia uma resposta de fuga.
Nota: A partir de nossa experiência, ~ 55% dos ratos será executado fora da plataforma central (sem rotação) enquanto a parte externa ainda está a rodar, e estes ensaios devem ser excluídos da análise. - Limpe a plataforma e abrigo e retornar o mouse para sua casa-gaiola, conforme descrito na etapa 4.10 e 4.11.
6. «ambiente dinâmico' ensaio comportamental:
Nota: Este ensaio destina-se para avaliar como os ratos se adaptar seus comportamentos defensivos para mudanças súbitas no ambiente. Nesses experimentos, após obter resposta de pelo menos uma fuga, o local do abrigo é alterado e o mouse está autorizado a visitá-lo uma vez antes de uma fuga subsequente resposta é disparada. Se o mouse perfeitamente atualiza a nova localização do abrigo e calcula a ausência de abrigo no seu local anterior, ela deve escapar para o novo local. Caso contrário, ou não memorizou o novo local do abrigo ou ele memorizou os dois locais, mas prefere fugir para o outro.
- Execute este ensaio depois passos 4.1 a 4.8 descrito acima, provocando pelo menos uma resposta de voo como um controle.
- Uma vez que o mouse voluntariamente sai do abrigo, remover o abrigo e coloque-o novamente em um novo buraco no extremo oposto da plataforma.
- Permitir que o mouse para explorar o ambiente até que entra o abrigo na sua nova localização.
- Quando o mouse sai do abrigo, presente com um estímulo contrário voluntariamente e observar sua resposta.
- Use o estímulo auditivo para este experimento por causa de sua longa duração, como sob a ameaça prolongada, animais podem executar respostas adicionais fuga para outros locais se eles não conseguem encontrar o abrigo em primeiro lugar.
- Limpe a plataforma e abrigo e retornar o mouse para sua casa-gaiola, conforme descrito nas etapas 4.10 e 4.11.
7. «sem abrigo» ensaio comportamental:
Nota: O objectivo deste teste é entender como a ausência de um abrigo no ambiente afeta a expressão de comportamentos defensivos.
- Conecte todos os 20 buracos. Neste experimento, há nenhum abrigo disponível na arena.
- Após a limpeza e secagem da plataforma (etapas 4.1 e 4.2), realizar etapas 4.4-4.6.
- Permita um período de habituação de 7 min.
- Apresentar o estímulo visual e observar a resposta defensiva. A falta de um abrigo promove o comportamento congelar, dura geralmente para a duração do estímulo visual. Use um ponto lentamente em expansão (ex. um círculo que subtende um ângulo visual de 2,6 ° pelo aparecimento e expande-se linearmente em 11,2 ° /s sobre 4 s para 47,4 °, no qual permanece por 1 s antes de deslocamento) para facilitar a observação de congelação respostas.
- Se mais de uma resposta por rato for necessária, use um estímulo inter intervalo pelo menos 60 s.
- Depois de obter o número necessário de respostas na ausência do abrigo, introduzir o abrigo sob um dos furos e deixe o mouse explorar o ambiente pelo menos 5 min. 5 min será tempo suficiente para a maioria dos ratos localizar e inserir o abrigo recém-introduzidos , mas caso isso não aconteça, alargar este período adicional 5 min blocos até pelo menos uma entrada é verificada.
- Repita as etapas de 4.8 e 4.9 e observar as respostas de fuga realizadas na presença de um abrigo no ambiente.
- Limpe a plataforma e abrigo e retornar o mouse para sua casa-gaiola, conforme descrito nas etapas 4.10 e 4.11.
8. os dados análise:
Nota: Cada uma das etapas de análise de dados a seguir pode ser executada independentemente.
- Respostas de fuga podem ser facilmente identificadas como uma aceleração súbita após a apresentação de um estímulo contrário. Quantificar a latência de fuga como o tempo entre a apresentação do estímulo e o início de um movimento de rotação de cabeça direcionado para o alvo da fuga ou o início do movimento de aceleração, o que vier primeiro.
- Congelamento de respostas pode ser caracterizado por total imobilidade do animal exceto respirando movimentos25, duradoura, pelo menos 0,5 s7. Medi a latência para congelar como o tempo entre a apresentação do estímulo contrário e o início do período de imobilidade.
- Para cada resposta de fuga, quantificar a linearidade da trajectória de voo calculando a razão entre a distância entre o rato e o abrigo e o deslocamento real durante o voo.
- Para cada resposta de fuga, calcule a precisão, medindo a distância entre onde o animal dirigido seu voo para o buraco e o buraco, levando para o abrigo. Uma vez que existem 20 furos, cada buraco fora do alvo representa um decréscimo de 10% na exatidão (ex., 2 furos fora do alvo podem ser expressa como 80% de precisão)
- Para o ensaio comportamental 'estratégia de navegação', quantificar a exatidão da resposta em relação ao local de abrigo antes da rotação da plataforma. Uma precisão média que não é significativamente diferente da precisão da pre-rotação indica que o animal não esteja utilizando qualquer uma das indicações sensoriais que foram giradas para navegar para o destino de fuga.
- Para o ensaio de 'ambiente dinâmico' comportamental, classifica cada resposta de fuga suscitou como sendo dirigido para o novo ou o antigo local de abrigo, ou para nenhum deles. Avalie como o padrão evolui ao longo as iterações dos comportamentos defensivos eliciadas.
- Para o ensaio comportamental 'sem abrigo', comparar a velocidade do mouse, após a apresentação do estímulo visual contrário na presença e na ausência de um abrigo e calcular a latência para o início da resposta defensiva. Além disso, calcule a probabilidade de desencadear uma resposta de congelamento em resposta ao estímulo para cada situação.
Resultados
Os ratos expostos a estímulos auditivos ou visuais iniciaram respostas de fuga rápida com latências curtas entre o início da estimulação e o início do voo. A latência média de escapar o estímulo visual foi 202 ± 16 ms (n = 51 respostas de 26 animais) e significativamente mais para estimulação auditiva: 510 ± 61 ms (n = 36 respostas de 15 animais, p< 0,0001 teste t entre visual e auditivo estimulação; Figura 2A) (passo 8.1). Respostas de fuga foram direcionadas com precisão para o abrigo (quer dizer precisão para estímulo visual: 97,2 ± 1,4% e 95,0 ± 1,4% para estímulo auditivo, não significativamente diferente entre os dois tipos de estímulos, p = 0.1655 t-teste; Figura 2B) (etapa 8,4), e as trajetórias de voo estavam perto de uma linha reta (proporção média de deslocamento com distância-abrigo para estímulo visual 107,3 ± 1,3% e 113.9 ± 1,5% para estímulo auditivo; A Figura 2) (passo 8.3). Observando uma curta latência do estímulo é importante distinguir evocada por estímulo do comportamento de homing executa, e a alta linearidade da trajetória do voo demonstra que a resposta de fuga tem o objetivo de alcançar o abrigo e não simplesmente se afastando desde o estímulo.
Apresentação de estímulo após a rotação da parte externa do labirinto enquanto o rato estava parado na plataforma central (intervalo = média de 36° - 90°, 56°) com êxito suscitou respostas de fuga (passo 5). Todos os ratos fugiram para o local anterior do abrigo, sem diminuição na precisão ou linearidade da trajetória de voo em comparação com os ensaios antes de rotação. A precisão média, medida em referência a posição original do abrigo, foi 96,3 ± 1,3% antes da rotação (n = 25 respostas de 8 ratos) e até mesmo um pouco maior após a rotação (100 ± 0%, p = 0,009 teste t entre pré e pós a rotação). A linearidade média em relação a localização original do abrigo foi 109.4 ± 5,0% pre-rotação (25 respostas de 8 ratos) e 109,1 ± 2,1%, pós-rotação (n = 8 respostas dos 8 ratos), p = 0.957 t-teste entre pré e pós-rotação, Figura 3A e B). Estes resultados demonstram que proximal visuais não são utilizados para orientar o voo para o abrigo (passos 8.3-8.5).
Mudar a localização do abrigo, depois de suscitar uma resposta de fuga bem sucedida (etapa 6) resultou em ratos a visitar o novo local de abrigo logo após a mudança (tempo médio para visitar o novo abrigo = 33,1 s, gama = s 4-82). Nos processos sequenciais de estimulação sonora produziram um aumento gradual da probabilidade de escapar para o novo local (44,4% no primeiro julgamento), atingindo 100% em todos os animais pelo quinto julgamento (n = 9 animais, Figura 3). Estes experimentos sonda a dinâmica de atualização da memória de local de abrigo para se adaptar às mudanças no ambiente (etapa 8,6).
Após um período de aclimatação de 7 min para o labirinto com sem abrigo (passo 7), a apresentação de um estímulo de tempo visual s 5 produzido comportamento congelar, ao invés de fuga (congelamento probabilidade = 95,2%, média de tempo de congelamento = 7,9 ± 2,7 s, n = 7 animais). Introdução posterior de um abrigo na mesma sessão e a apresentação do estímulo visual tempo suscitou escapar respostas confiável, mostrando que os ratos com flexibilidade podem trocar estratégias defensivas, dependendo da presença de refúgio (Figura 3D) ( etapas 8.1 e 8.7).

Figura 1: esquemático da arena comportamental. Desenho dos componentes principais da arena comportamental, visto de cima (à direita) e do lado (à esquerda). Uma grande fração da plataforma pode ser girada, incluindo o abrigo e indicações visuais anexadas. Olheiras na vista top são os buracos que o abrigo pode ser anexado a, que são todos idênticos (um exemplo de local de abrigo é indicado). A arena inteira está contida em um gabinete de amortecimento de som. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: respostas de fuga são rápidos e precisos comportamentos meta-direcionado. (A) a latência para o início da fuga desde o início da estimulação visual ou auditivo. (B) a precisão da navegação para o abrigo durante a fuga, medida tal que o valor é 100% se o rato corre diretamente para o abrigo e 10% menos para cada buraco longe do abrigo (EG., 80% se o buraco alcançado primeiro foi dois furos longe de uma correta). (C) deslocamento do mouse durante fuga plotada contra a distância linear entre a posição inicial do animal e o local de destino, mostrando que o voo está perto de linear. Caixa e bigodes mostram dados de percentil 10-90, e os restantes pontos de dados são mostrados como círculos. Os gráficos de cor vermelha são dados de estimulação auditiva (n = 36 respostas de 15 animais) e preto são de estimulação visual (n = 51 respostas de 26 animais). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: ratos não usar pistas visuais proximais ou sugestões relacionados ao abrigo para orientar respostas de fuga. (A) enredo de deslocamento como na Figura 2, para as respostas pré e pós-rotação da plataforma. (B) a precisão da navegação durante fuga em condições pré e pós-rotação. Precisão pós-rotação é medida em referência a localização inicial do abrigo e mostra que os ratos navegar de volta para o local original do abrigo em vez da nova posição. (C) fração de respostas de fuga, direcionadas para o novo local de abrigo, plotado contra número de estimulação. Após a apresentação deth 5 do estímulo, todos os animais escapar diretamente para o novo local de abrigo (n = 9 animais). (D) significa velocidade do outro lado os animais em resposta a estímulos visuais, com um abrigo disponível (vermelho) ou ausente (azul), mostrando a fuga e congelamento respostas, respectivamente. Velocidade foi normalizada para a linha de base e alinhada ao tempo de reação (definido como também o início de um voo ou resposta de congelamento, linha tracejada). Áreas sombreadas mostram SEM. (n = 7 animais). Para 2A e 2B, a caixa e bigodes mostrar dados de percentil 10-90, e os restantes pontos de dados são mostrados como círculos. Os gráficos de cor vermelha são dados de estimulação auditiva antes de rotação da plataforma (n = 25 respostas de 8 animais) e azul são de respostas depois de girar a plataforma (n = 8 respostas de 8 animais). Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Os ensaios que descrevemos aqui são tecnicamente simples de executar, e com excepção da plataforma experimentos de rotação, pode ser implementado facilmente em um labirinto de Barnes padrão. No entanto, algumas complicações podem surgir: por um lado, o rato pode parecer com medo de entrar no abrigo, que pode ser devido ao abrigo de não ser suficientemente limpos ou secas. Por outro lado, o mouse pode ficar dentro do abrigo por longos períodos de tempo. É crítico para estas experiências que os animais são nunca removidos do abrigo manualmente, como isso pode perturbar a integração do caminho, bem como a angústia dos animais e mudar sua resposta à ameaça. Sugerimos que encerra o experimento após 90 min e refazer o teste após 48 h se necessário. Uma outra consideração prática relevante é que quando o mouse investiga a borda da plataforma, não pode detectar estímulos visuais se sua cabeça é dobrada sobre a borda, e, portanto, é recomendável reter os estímulos em tais situações. Além disso, os ratos podem saltar fora da plataforma em raras ocasiões. Sugerimos que terminando o experimento e refazer o teste após 48 h. ratos que ativamente deixar a plataforma uma vez poderão fazê-lo novamente no futuro e podem ter que ser excluídos do estudo. Finalmente, de nossa experiência, a maioria dos ratos responderá ao estímulo visual (34/36 ratos). No entanto, se os ratos não mostram qualquer tipo de resposta ao estímulo visual (um alarme, um voo ou uma resposta de congelamento), eles devem ser excluídos do estudo.
Um aspecto importante a considerar é que o controle de sinais sensoriais normalmente é um problema em ensaios de memória espacial, como muitas vezes é difícil de eliminar todas as possíveis fontes de contaminação pistas. Nossa configuração experimental reduz visuais externas, em torno do labirinto com uma parede, e é colocado dentro de um armário de amortecimento-som para isolamento acústico. Para minimizar as sugestões olfactory, recomendamos limpar cuidadosamente a afinação com 70% de etanol ou ácido acético entre experiências.
Nosso ensaio comportamental adiciona métodos anteriores para estudar navegação espacial26 usando comportamentos defensivos instintivos para sondar a memória espacial e com foco em cenários etológicos. Uma diferença fundamental entre os procedimentos descritos aqui e o teste padrão do BM é a falta de sessões de treinamento. Em nossos ensaios, o período de habituação garante que o mouse visita o abrigo pelo menos uma vez e, frequentemente, muitas vezes mais, que temos demonstrado anteriormente para ser suficientes para memorizar o abrigo local2. Acreditamos que uma razão importante para a elevada taxa de sucesso e precisão em encontrar o abrigo apesar da falta de treinamento nunca passivamente é deslocar o animal durante o experimento, tornando assim a integração do caminho uma estratégia viável de navegação. No entanto notamos que nosso ensaio lida com memória formada e avaliados durante uma única sessão, e que nós não testamos para memória de longo prazo da localização do abrigo, que geralmente é o objetivo do padrão BM experiências. Finalmente, usando estímulos intrinsecamente contrário discretos ao invés do fã comumente usado ou buzz para a duração da sessão, nosso ensaio fornece controle experimental muito bom em dois comportamentos que podem usar estratégias diferentes de navegação: forrageamento e fuga de abrigo-dirigido. Acreditamos que o uso destes ensaios em combinação com técnicas da moderna neurociência para gravação e manipulação de atividade neural pode fornecer insights importantes sobre circuitos neurais como calcular o comportamento.
Divulgações
A maioria dos dados incluídos na seção de resultados é um subconjunto dos dados apresentados no Vale et. al 2017.
Agradecimentos
Este trabalho foi financiado por um Wellcome Trust/real sociedade Henry Dale Fellowship (098400/Z/Z/12), um Conselho de pesquisa médica (MRC) conceder MC-UP-1201/1, Wellcome Trust e Gatsby Charitable Foundation SWC Fellowship (para tuberculose), MRC PhD Studentship (D.E. e RV) e uma bolsa de doutoramento de Fonds Boehringer Ingelheim (para RV). Agradecemos a Kostas Betsios para o software de aquisição de dados, o LMB mecânicos e elétricos oficinas para construção da arena experimental de programação.
Materiais
| Name | Company | Catalog Number | Comments |
| Infrared iluminators TV6700 | Abus | - | |
| DLP projector Infocus IN3126 | Infocus | 0001740992-00000001 | |
| Ultrasound speaker Pettersson L60 | Pettersson Elektronik | - | |
| Amplifier QTX PRO240 | QTX | - | |
| Soundcard Xonar D2 | Asus | 90-YAA021-1UAN00Z | |
| HP Z840 desktop | HP | F5G73AV | |
| 100 micron drafting film | Xerox | 3R98145 | |
| Near infrared camera: Basler acA1300-60gmNIR | Basler | 106202 | |
| National Instruments BNC-2110 | National instruments | 777643-01 | |
| LabVIEW 2015 64-bit | National instruments | - | |
| Custom made Barnes maze | MRC Laboratory of Molecular Biology mechanical workshop | - |
Referências
- Eaton, R. C., Bombardieri, R. A., Meyer, D. L. The Mauthner-initiated startle response in teleost fish. J Exp Biol. 66 (1), 65-81 (1977).
- Vale, R., Evans, D. A., Branco, T. Rapid Spatial Learning Controls Instinctive Defensive Behavior in Mice. Curr Biol. 27 (9), 1342-1349 (2017).
- Blanchard, R. J., Flannelly, K. J., Blanchard, D. C. Defensive behavior of laboratory and wild Rattus norvegicus. J Comp Psychol. 100 (2), 101-107 (1986).
- Dill, L. M., Houtman, R. The influence of distance to refuge on flight initiation distance in the gray squirrel (Sciurus carolinensis). Canadian Journal of Zoology. 67 (1), 233-235 (1989).
- Blanchard, R. J., Blanchard, D. C., Rodgers, J., Weiss, S. M. The characterization and modelling of antipredator defensive behavior. Neurosci Biobehav Rev. 14 (4), 463-472 (1990).
- Fanselow, M. S., Lester, L. S. A functional behavioristic approach to aversively motivated behavior: Predatory imminence as a determinant of the topography of defensive behavior. Evolution and learning. , (1988).
- De Franceschi, G., Vivattanasarn, T., Saleem, A. B., Solomon, S. G. Vision Guides Selection of Freeze or Flight Defense Strategies in Mice. Curr Biol. 26 (16), 2150-2154 (2016).
- Bräcker, L. B., et al. Essential role of the mushroom body in context-dependent CO 2 avoidance in Drosophila. Current Biology. 23 (13), 1228-1234 (2013).
- Ghosh, D. D., et al. Neural architecture of hunger-dependent multisensory decision making in C. elegans. Neuron. 92 (5), 1049-1062 (2016).
- Bellman, K. L., Krasne, F. B. Adaptive complexity of interactions between feeding and escape in crayfish. Science. 221 (4612), 779-781 (1983).
- Rodgers, W., Melzack, R., Segal, J. "Tail flip response" in goldfish. Journal of comparative and physiological psychology. 56 (5), 917(1963).
- Dill, L. M. The escape response of the zebra danio (Brachydanio rerio) II. The effect of experience. Animal Behaviour. 22 (3), 723-730 (1974).
- Barnes, C. A. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 93 (1), 74-104 (1979).
- Pompl, P. N., Mullan, M. J., Bjugstad, K., Arendash, G. W. Adaptation of the circular platform spatial memory task for mice: use in detecting cognitive impairment in the APP(SW) transgenic mouse model for Alzheimer's disease. J Neurosci Methods. 87 (1), 87-95 (1999).
- Patil, S. S., Sunyer, B., Höger, H., Lubec, G. Evaluation of spatial memory of C57BL/6J and CD1 mice in the Barnes maze, the Multiple T-maze and in the Morris water maze. Behavioural Brain Research. 198 (1), 58-68 (2009).
- Sunyer, B., Patil, S., Höger, H., Lubec, G. Barnes maze, a useful task to assess spatial reference memory in the mice. Protocol Exchange. , (2007).
- Attar, A., et al. A shortened Barnes maze protocol reveals memory deficits at 4-months of age in the triple-transgenic mouse model of Alzheimer's disease. PLoS One. 8 (11), e80355(2013).
- Harrison, F. E., Reiserer, R. S., Tomarken, A. J., McDonald, M. P. Spatial and nonspatial escape strategies in the Barnes maze. Learn Mem. 13 (6), 809-819 (2006).
- Leary, T. P., Brown, R. E. The effects of apparatus design and test procedure on learning and memory performance of C57BL/6J mice on the Barnes maze. J Neurosci Methods. 203 (2), 315-324 (2012).
- Mittelstaedt, M. -L., Mittelstaedt, H. Homing by path integration in a mammal. Naturwissenschaften. 67 (11), 566-567 (1980).
- Stackman, R. W., Golob, E. J., Bassett, J. P., Taube, J. S. Passive transport disrupts directional path integration by rat head direction cells. J Neurophysiol. 90 (5), 2862-2874 (2003).
- Yilmaz, M., Meister, M. Rapid innate defensive responses of mice to looming visual stimuli. Curr Biol. 23 (20), 2011-2015 (2013).
- Mongeau, R., Miller, G. A., Chiang, E., Anderson, D. J. Neural correlates of competing fear behaviors evoked by an innately aversive stimulus. J Neurosci. 23 (9), 3855-3868 (2003).
- Hurst, J. L., West, R. S. Taming anxiety in laboratory mice. Nat Methods. 7 (10), 825-826 (2010).
- Paylor, R., Tracy, R., Wehner, J., Rudy, J. W. DBA/2 and C57BL/6 mice differ in contextual fear but not auditory fear conditioning. Behavioral neuroscience. 108 (4), 810(1994).
- Paul, C. M., Magda, G., Abel, S. Spatial memory: Theoretical basis and comparative review on experimental methods in rodents. Behav Brain Res. 203 (2), 151-164 (2009).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados