
Method Article
Un ensayo conductual para investigar el papel de la memoria espacial durante defensa instintiva en ratones
En este artículo
Resumen
Este protocolo describe un ensayo conductual, basado en la prueba del laberinto de Barnes, para estudiar cómo instintiva defensiva acciones son modificadas por el conocimiento del entorno espacial.
Resumen
Evolución ha seleccionado un repertorio de comportamientos defensivos que son esenciales para la supervivencia a través de todas las especies animales. Estos comportamientos son a menudo estereotipados acciones provocadas en respuesta a estímulos sensoriales de forma innata aversión, pero su éxito requiere de suficiente flexibilidad para adaptarse a diferentes ambientes espaciales, que pueden cambiar rápidamente. Aquí, describimos un análisis conductual para evaluar la influencia del conocimiento espacial aprendido comportamientos defensivos en ratones. Hemos adaptado el ensayo utilizado de memoria espacial de laberinto Barnes para investigar cómo ratones vaya a un refugio durante las respuestas de escape de forma innata aversión estímulos sensoriales en un ambiente de novela, y cómo se adaptan a los cambios agudos en el entorno. Este nuevo ensayo es un paradigma etológico que no requiere de formación y aprovecha los patrones de exploración natural y estrategias de navegación en ratones. Proponemos que el conjunto de protocolos descritos aquí son un medio poderoso de estudiar conductas intencionadas y navegación activada por el estímulo, que debe ser de interés para los campos de conductas instintivas y memoria espacial.
Introducción
Conductas instintivas de defensa en gran parte se consideran ser hardwired estímulo-respuesta, como el movimiento de arranque C en peces y anfibios, que se mueve el animal lejos de una fuente de amenaza1. Sin embargo, conductas defensivas pueden ser más adaptativos si flexibilidad toman en cuenta la información aprendida sobre el entorno actual. Un ejemplo de esta flexibilidad es el cambio de escape a congelación se muestra por los roedores cuando amenaza, dependiendo del conocimiento previo sobre la presencia o ausencia de un refugio en el medio ambiente2,3. Otros ejemplos de flexibilidad en los comportamientos naturales incluyen la adaptación del umbral de iniciación de vuelo o escapan de velocidad dependiendo de la distancia entre una presa y su refugio4, distancia amenaza5,6y anterior experiencia 11 , 12, así como seleccionar diferentes estrategias defensivas dependiendo de las propiedades sensoriales del estímulo aversivo7, o incluso suprimir el comportamiento defensivo frente a competidores motivaciones tales como hambre8, 9,10. La dependencia de selección de acción en el conocimiento adquirido acerca de las características espaciales del medio ambiente hace conductas instintivas de defensa en ratones un poderoso modelo para el estudio de selección de objetivo, memoria espacial y la navegación. Aquí, describimos la adaptación de una tarea conductual utilizada, análisis de la memoria espacial del laberinto (BM) Barnes13, para determinar la influencia del ambiente espacial en la selección de la acción defensiva en los ratones y sus estrategias de navegación cuando escapar hacia un refugio.
El laberinto de Barnes estándar utilizado para estudiar el aprendizaje memoria espacial en ratones consiste en una plataforma de diámetro circular ~ 90 cm con 20 orificios equidistantes, de las cuales 19 están cerrados y uno conduce a un refugio subterráneo que ratones buscan evitar el campo abierto para entorno de la plataforma. Débiles estímulos aversivos (timbre, luz, ventilador) se utilizan a menudo, continuamente a lo largo del ensayo para hacer el entorno aversivo y así promover la entrada en el refugio14. En el ensayo más utilizado15,16, el animal tiene un ensayo de habituación donde es guiado hasta el refugio manualmente por el experimentador inmediatamente después de ser colocado en la plataforma. Esto es seguido por un período de 4 días de adquisición, donde cada día que el ratón puedo navegar libremente en el laberinto durante 3 min, después de que es otra vez manualmente guiado al refugio si no alcanza durante el período de exploración. La última etapa del análisis es una sonda de prueba en la 5º jornada (aunque también comúnmente se realiza una memoria a largo plazo de siete días prueba) cuando el animal explora el laberinto con todos los orificios cerrados. Memoria aprendizaje y a largo plazo se cuantifican por el tiempo necesario para encontrar el refugio y pinchazos en los agujeros equivocados durante el período de adquisición y por el tiempo cerca del agujero cerrado refugio en la prueba de la sonda. Resultados típicos muestran una disminución en el número de errores y la latencia para alcanzar el refugio durante la adquisición y una proporción por encima de la posibilidad de tiempo en el cuadrante que contiene el orificio cerrado blanco en la punta de prueba prueba15.
Mientras que varias variantes del ensayo BM han sido previamente descritos17,18,19, el paradigma que Describimos aquí tiene tres cambios fundamentales desde el ensayo estándar. En primer lugar, el animal quedo por explorar el laberinto y encontrar el refugio en su propia, y la prueba es hecha en la misma sesión, poco después de que se ha encontrado el refugio durante los períodos donde el animal se dedica a la conducta exploratoria. Aunque esta opción no comprueba memoria a largo plazo de la situación de refugio, se diseña un escenario naturalista que imita a la exploración del nuevo territorio, bajo la amenaza de depredación. Además, permite realizar pruebas de cómo los animales se adaptan a los cambios agudos en el entorno, como cambios bruscos de paisaje. En segundo lugar, un aspecto clave de nuestro análisis es que el experimentador no fuerzas el animal dentro o fuera de la vivienda, que puede desorientar al animal y excluir la ruta de integración20 como una estrategia viable de la navegación del21. Camino de la integración es una estrategia de navegación que utiliza señales de uno mismo-movimiento, tales como señales propioceptivas y vestibulares, derivados de la integración del motor de la salida, para actualizar la posición actual del animal y navegar hacia una meta, que no es posible si el animal se mueve pasivamente por el experimentador. En tercer lugar, utilizamos visual innata aversión22 y estímulos auditivos23 para escapar a la vivienda, que se distingue fácilmente del comportamiento de alimentación continua y permite la evaluación y cuantificación de estrategias específicas de navegación durante la defensa de las amenazas inminentes. Proponemos que este análisis será útil para diseccionar el papel de la memoria espacial en la selección y aplicación de conductas defensivas, y más generalmente en el más amplio estudian de meta-dirigida de la navegación y la memoria espacial a corto plazo. Los protocolos descritos aquí fueron introducidos por Vale et al. en 2017, a que nos referimos a los lectores más en discusión de la profundidad de los fundamentos para los experimentos y resultados.
Protocolo
Todos los experimentos se realizaron bajo la ley de Reino Unido los animales (procedimientos científicos) de 1986 (PPL 70/7652) tras la aprobación ética local.
1. puesta en funcionamiento del aparato conductual
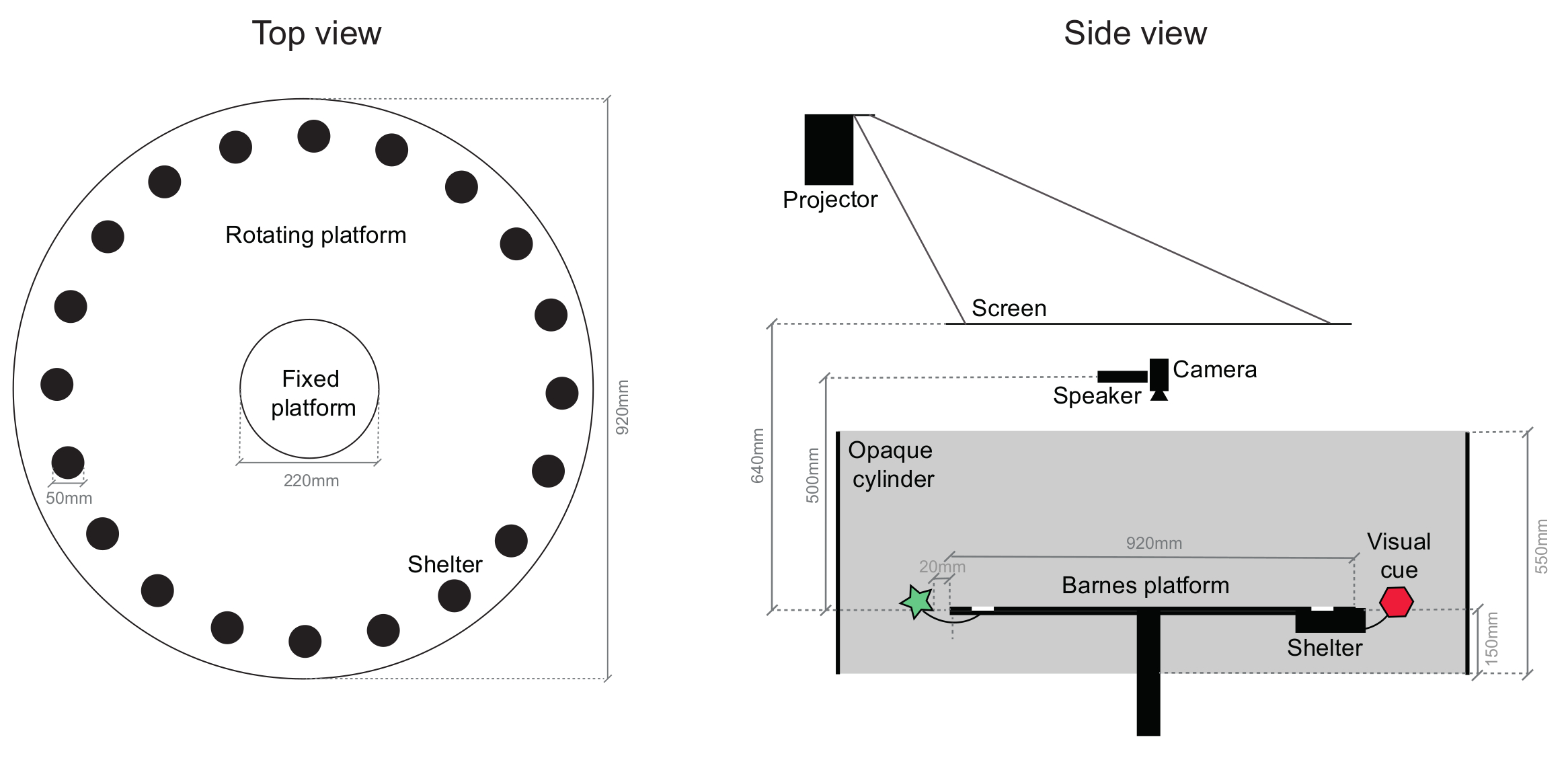
Nota: El principal componente del aparato del comportamiento es una variante del laberinto de Barnes13 consisten en una plataforma circular de acrílico blanco (92 cm de diámetro) con equidistante de 20, 5 cm de diámetro, orificios circulares, ubicados radialmente, 5 cm desde el borde de la plataforma . 19 de estos son para cerrar con tapón de plástico negro (5 mm de profundidad), mientras que el agujero restante conduce a un refugio de metacrilato negro (dimensiones 15 x 5,8 x 4,7 cm) que ratones fácilmente pueden entrar y salir. La zona central de la arena consiste en una plataforma circular fija (22 cm de diámetro) y la periferia se monta en un marco que permite una rotación más de 360 grados a través de un motor de pasos controlado por ordenador. La plataforma está elevada 45 cm desde el piso para evitar que los ratones bajando. Ver dibujo esquemático en la figura 1.
- Conectar visual de señales a la periferia de la plataforma (2D o 3D símbolos con diferentes formas, colores y patrones), por lo menos 2 cm del borde para evitar que ratones alcanzarlas.
Nota: Los ejemplos de señales visuales son cuadrados azules llano de 262 cm2, o círculos (de 15,5 cm de diámetro) con franjas diagonales de color púrpura. - Añadir ~ 2 g del lecho de la jaula del ratón casero al interior de la vivienda para servir como una señal olfativa local. Opcionalmente, colocar una luz de diodo (LED) en la entrada de la vivienda para proporcionar una indicación de señal visual local.
- Para aislar el ratón de externos, señales visuales distales, montar una pared octogonal o cilíndrica, extensible de 40 cm y 15 cm bajo la superficie de la plataforma, alrededor de la arena, coloca 10 cm de distancia del borde de la plataforma.
- Coloque un altavoz ultrasonido 50 cm encima del centro de la plataforma, para ofrecer estímulos auditivos aversivos.
- Coloque una pantalla translúcida 64 cm por encima de la arena a la espalda-proyecto arriba aversión estímulos visuales usando un proyector.
- Proporcionada por el proyector se encenderá el aparato conductual; calibrar por lo que hay 5-10 lx en el centro de la superficie de la plataforma. Coloque luces infrarrojas y una cámara infrarroja sobre la plataforma, con un filtro de paso largo (> 700 nm) para prevenir el parpadeo del proyector en el video adquirido.
- Seguimiento de la posición del ratón en línea, utilizando el software disponible comercialmente o un algoritmo adecuado. Para el seguimiento ratones negro en un blanco de la arena, una operación simple umbral seguida por cálculo de centroide de la máscara es apropiada.
- Idealmente, coloque el aparato de comportamiento dentro de una caja de prueba de sonido y luz, para evitar que las señales externas que afectan el experimento.
2. entrega de estímulos aversivos innatamente
Nota: Los estímulos que se describe a continuación pueden ser generados con una amplia gama de paquetes de software diferentes, como LabVIEW y Matlab.
- Estímulo visual
- Proyecto de los estímulos en la pantalla colocada 64 cm sobre la arena. El estímulo consiste en un forma lineal creciente círculo oscuro sobre un fondo gris22 (contraste de Weber =-0.98, luminancia = 7,95 cd/m2).
Nota: El círculo estándar ideal debe estar centrado directamente sobre la posición del ratón, abrazadas un ángulo visual de 2,6 ° al inicio y se expande linealmente en 224 ° /s más 200 ms a 47,4 °, en el cual sigue siendo de 250 ms antes de offset. - Para entregar el estímulo visual, centro sobre el ratón. Esto puede lograrse por una espera hasta que el ratón pasa a través de una región previamente definida en la arena, o siguiendo la posición del ratón en línea y usando las coordenadas de seguimiento para definir el centro del círculo.
Nota: Aunque sugerimos proyectar el estímulo visual directamente sobre el ratón, comportamientos defensivos pueden también ser sacados si el mismo estímulo se proyecta por delante del ratón (aunque todavía paralelos al suelo). En nuestra experiencia, este es el caso por lo menos hasta 30 cm por delante de la posición del ratón.
- Proyecto de los estímulos en la pantalla colocada 64 cm sobre la arena. El estímulo consiste en un forma lineal creciente círculo oscuro sobre un fondo gris22 (contraste de Weber =-0.98, luminancia = 7,95 cd/m2).
- Estímulo auditivo
- Desencadenar un estímulo que consiste en un tren de tres frecuencia modulada hasta-barridos de 17 a 20 kHz23 3 s, durando 9 s en total. El nivel de presión acústica (SPL), medido a nivel de la plataforma, debe ser en un rango de entre 73 y 78 dB.
- Conecte una tarjeta de sonido y un amplificador al altavoz ultrasonido colocado sobre la arena para entregar el estímulo auditivo.
3. los animales:
- Utilizar ratones C57BL/6J hombres o mujeres, entre 6 y 24 semanas de edad. Mantener que los animales en una luz de 12 h ciclo con acceso gratuito a chow y agua.
- Asegurar que los ratones solo ocupan al menos 72 h antes de la prueba y del análisis conductual durante la fase de luz de la luz ciclo.
Nota: Encontramos que el comportamiento exploratorio inicial de ratones alojados solo es más comparable a través de animales que en ratones alojados en grupo. Además, animales con implantes quirúrgicos para registro de actividad de los nervios o manipulación a menudo necesitan ser único ubicado, y por lo tanto puede ser beneficioso tener conjuntos de datos de control en las mismas condiciones de la vivienda.
4. estándar análisis conductual:
- Limpiar meticulosamente la superficie de la plataforma con etanol al 70% o con ácido acético para eliminar señales olfativas no deseados. El refugio con agua de enjuague y lave con etanol al 70%, después de lo cual debería ser enjuagado con agua para disminuir el olor del etanol.
- Seque la plataforma y el refugio, como el ratón puede evitar entrar en el refugio si está mojado.
- Aleatoria la ubicación de la vivienda en la plataforma de cada ensayo girando la plataforma antes del experimento. La ubicación del refugio será anclada en la arena pero se giran en relación a la caja del aparato conductual.
- Llevar el ratón a la sala de experimento en su casa-jaula y colocar la jaula sobre la plataforma de prueba por un período de aclimatación de 10 minutos.
- Quitar el ratón de su jaula de la casa y evitar recuperar la cola: lo de la taza o dejarlo subirse a un elemento de enriquecimiento que se puede levantar. Ratones por la cola ha sido mostrar a aumentar los niveles de ansiedad y así pueden afectar las respuestas del comportamiento para amenaza24.
- Suavemente Coloque el ratón en el centro de la arena y empezar la adquisición de video.
- Permitir un periodo de habituación de 7 min, durante el cual ningún estímulo aversivo se presenta. Asegúrese de que el ratón tiene tiempo suficiente para encontrar y entrar en el refugio por lo menos una vez. Lo consideran como un ingreso si las cuatro extremidades están dentro de la vivienda. 7 minutos debería ser suficiente para la mayoría de los ratones para encontrar y entrar en el refugio, pero en caso de que esto no suceda, extender el período de habituación por períodos adicionales de 5 minutos hasta que se haya verificado al menos una entrada.
- Activar la entrega de los estímulos aversivos (auditiva o visual) manualmente observando el video en vivo, o automáticamente por la posición del animal en línea de seguimiento y pre-definir una región de interés en la arena para el disparo de estímulos-entrega una vez que el animal alcanza. El estímulo visual se entrega generalmente directamente sobre el animal mediante el video online seguimiento de coordenadas.
- Para obtener más de una respuesta de escape en la misma sesión, dejar el ratón voluntariamente el refugio. Si el ratón no se escaparon con éxito al refugio en el último estímulo aversivo, espere al menos 60 s antes de presentar otro estímulo.
- Una vez terminado el ensayo, regresar el animal a su jaula casa. En caso de que se requiere más pruebas del mismo ratón, espere al menos 48 h antes de repetir el análisis.
- Limpiar la plataforma y el refugio como se describe en el paso 4.1 y 4.2 antes de probar el siguiente animal.
Nota: Los pasos 5, 6 y 7 son independientes del análisis conductual estándar y cada uno puede realizar de forma individual.
5 ' estrategia de navegación' análisis conductual:
Nota: El objetivo de este ensayo es determinar qué señales ratones usan para guiar el comportamiento del escape. En este ensayo, se gira la parte móvil externa de la plataforma cuando el ratón está en el centro de la arena fija, que se desplaza radialmente el refugio, las señales olfativas ubicadas dentro del refugio y el proximal señales visuales a la plataforma. Si el ratón está siguiendo cualquiera de las señales que han sido giradas para navegar hasta el refugio, correctamente se escapará a la nueva ubicación de la vivienda, pero si utiliza cualquier otras señales que han permanecido en su lugar, se vaya a la ubicación anterior del refugio.
- Realice los pasos 4.1 a 4.8 de la prueba de comportamiento estándar descrito anteriormente, suscitar al menos una respuesta de huida para ser utilizado como línea de base.
- Deja el ratón activamente dejar el refugio y esperar a que vaya a la parte central de la plataforma. Coloque un plato pequeño con ropa de cama en el centro para fomentar la exploración y mantener el animal en la plataforma central lo suficientemente largos para permitir la rotación de la parte exterior de la plataforma. Añadir el plato y ropa de cama mientras el ratón está dentro de la vivienda y después de la exitosa fuga que precede a la rotación de la plataforma. Si el ratón no puede entrar en el refugio durante la respuesta de escape, espere a entrar voluntariamente en el refugio y luego añadir el plato y ropa de cama.
- Utilice el motor de pasos para girar la plataforma al menos 36 grados (2 agujeros) y luego usar inmediatamente un estímulo sensorial para provocar una respuesta de escape. Es posible que la rotación activa un susto e incluso inicia una respuesta de escape.
Nota: Por nuestra experiencia, ~ 55% de los ratones se ejecutará fuera de la plataforma central (no giratorio) mientras que la porción externa está girando, y estos ensayos deben ser excluidos del análisis. - Limpiar la plataforma y refugio y retomar el ratón de su jaula de inicio como se describe en el paso 4.10 y 4.11.
6. ' ambiente dinámico' análisis conductual:
Nota: Este ensayo está diseñado para evaluar cómo ratones adaptan sus comportamientos defensivos a cambios repentinos en el medio ambiente. En estos experimentos, después de que la respuesta de escape al menos una, se cambia la ubicación del refugio y el ratón se permite visitarlo una vez antes de un escape posterior se desencadena la respuesta. Si el ratón perfectamente actualiza la nueva ubicación de la vivienda y computa la ausencia del refugio en su ubicación anterior, debe escapar a la nueva ubicación. De lo contrario, no ha memorizado la nueva ubicación de la vivienda o ha memorizado ambas localidades pero prefiere escapar a la antigua.
- Realizar este ensayo después de pasos 4.1 a 4.8 describen anteriormente, que por lo menos una respuesta de huida como un control.
- Una vez que el ratón abandona voluntariamente el refugio, quite el abrigo y volver a colocarlo en un nuevo agujero en el extremo opuesto de la plataforma.
- Permitir que el ratón para explorar el ambiente hasta que entra en el refugio en su nueva ubicación.
- Cuando el ratón voluntariamente viene del refugio, presente con un estímulo aversivo y observar su respuesta.
- Utiliza el estímulo auditivo para este experimento de debido a su mayor duración, como bajo amenaza prolongada, animales pueden llevar a cabo las respuestas de escape adicional a otras localidades si no logran encontrar el refugio en un principio.
- Limpiar la plataforma y refugio y retomar el ratón de su jaula de inicio como se describe en los pasos 4.10 y 4.11.
7. ' no Shelter' análisis conductual:
Nota: El objetivo de este ensayo es entender cómo la ausencia de un refugio en el medio ambiente afecta a la expresión de comportamientos defensivos.
- Tapar todos los agujeros de 20. En este experimento, no se dispone de ningún abrigo en la arena.
- Después de la limpieza y secado de la plataforma (pasos 4.1 y 4.2), realizar pasos 4.4-4.6.
- Permite un periodo de habituación de 7 min.
- Presentar el estímulo visual y observar la respuesta defensiva. La falta de un refugio promueve el comportamiento congelación, generalmente dura para la duración del estímulo visual. Usar un spot lentamente creciente (ej. un círculo que subtienda un ángulo visual de 2,6 ° al inicio y se expande linealmente en 11.2 ° /s en 4 s a 47,4 °, en el cual permanece durante 1 s antes offset) para facilitar la observación de las respuestas de congelación.
- Si se requiere más de una respuesta por ratón, usar un intervalo inter-estímulo de al menos 60 s.
- Después de obtener el número requerido de respuestas en la ausencia de la vivienda, introducir el refugio debajo de uno de los agujeros y dejar el ratón explorar el medio ambiente durante al menos 5 min 5 min será tiempo suficiente para la mayoría Ratones encontrar y entrar en el refugio recién introducido , pero en caso de que esto no suceda, extender este período por 5 min más bloques hasta que se verifique al menos una entrada.
- Repita los pasos del 4.8 y 4.9 y observar las respuestas de escape realizadas en presencia de un refugio en el medio ambiente.
- Limpiar la plataforma y refugio y retomar el ratón de su jaula de inicio como se describe en los pasos 4.10 y 4.11.
8. Análisis:
Nota: Cada uno de los pasos de análisis de datos puede realizarse independientemente.
- Las respuestas de escape pueden ser fácilmente identificadas como una repentina aceleración después de la presentación de un estímulo aversivo. Cuantificar la latencia de escape como el tiempo entre la presentación del estímulo y el comienzo de un movimiento de rotación de la cabeza dirigido hacia el blanco del escape o el inicio del movimiento de aceleración, lo que ocurra primero.
- Respuestas de congelación se caracteriza por la total inmovilidad del animal salvo respirar movimientos25, duradero por lo menos 0,5 s7. Medir la latencia para congelar el tiempo entre la presentación del estímulo aversivo y el inicio del período de inmovilidad.
- Para cada respuesta de escape, cuantificar la linealidad de la trayectoria de vuelo calculando el cociente de la distancia entre el ratón y el refugio y el desplazamiento real durante el vuelo.
- Para cada respuesta de escape, calcular la exactitud midiendo la distancia entre el orificio donde el animal dirige su vuelo y el agujero que conduce al refugio. Puesto que hay 20 agujeros, cada hoyo de destino representa un decremento del 10% en la precisión (por ej., 2 agujeros de objetivo pueden ser expresados como 80% de precisión)
- Para el análisis del comportamiento de 'estrategia de navegación', cuantificar la exactitud de la respuesta en relación con la ubicación de la vivienda antes de la rotación de la plataforma. Una precisión promedio que no es significativamente diferente de la exactitud de la rotación previa indica que el animal no está usando ninguna de las señales sensoriales que se rotaban para navegar en el destino de escape.
- Para el análisis del comportamiento de 'ambiente dinámico', clasificar cada respuesta de escape provocada como ser dirigidas hacia la nueva o la vieja localización de vivienda, o a ninguno de ellos. Evaluar cómo evoluciona el patrón a lo largo de las iteraciones de las conductas defensivas sacadas.
- Para el análisis de comportamiento 'no shelter', comparar la velocidad del ratón después de la presentación del estímulo aversivo visual en presencia y en ausencia de un refugio y calcular la latencia a la iniciación de la respuesta defensiva. Además, calcular la probabilidad de desencadenar una respuesta de congelación en respuesta al estímulo para cada situación.
Resultados
Ratones expuestos a estímulos auditivos o visuales iniciaron las respuestas de escape rápido con poca latencia entre el inicio del estímulo y la iniciación del vuelo. La latencia media para escapar de los estímulos visuales fue 202 ± 16 ms (n = 51 respuestas de 26 animales) y significativamente mayor para la estimulación auditiva: 510 ± 61 ms (n = 36 respuestas de 15 animales, p< 0.0001 t-test entre visual y auditiva estimulación; Figura 2A) (paso 8.1). Las respuestas de escape fueron dirigidas con precisión al refugio (significa exactitud para estímulos visuales: 97,2 ± 1.4% y 95.0 ± 1.4% para estímulo auditivo, no significativamente diferente entre los dos tipos de estímulos, p = 0.1655 prueba t; Figura 2B) (paso 8.4), y trayectorias de vuelo cerca de una línea recta (ratio medio de desplazamiento a distancia al refugio para estímulos visuales ± 1,3% 107,3 y 113,9 ± 1,5% de estímulo auditivo; Figura 2) (paso 8.3). Observando un tiempo de latencia corto desde el estímulo es importante distinguir la conducta evocada por el estímulo de recalada funciona, y la alta linealidad de la trayectoria del vuelo demuestra que la respuesta de escape tiene el objetivo de alcanzar el refugio y no simplemente alejando desde el estímulo.
Presentación de estímulo después de la rotación de la parte exterior del laberinto mientras que el ratón estaba inmóvil en la plataforma del centro (rango = promedio de 36° - 90°, 56°) con éxito generó respuestas de escape (paso 5). Todos los ratones huyeron hacia la ubicación anterior de la vivienda, con ninguna disminución en la exactitud y linealidad de la trayectoria de vuelo en comparación con los ensayos antes de rotación. La precisión media, medida en referencia a la posición original de la vivienda, fue de 96,3 ± 1.3% antes de la rotación (n = 25 respuestas de 8 ratones) y un poco más altos después de la rotación (100 ± 0%, p = 0.009 t-test entre pre y post rotación). La linealidad de la media en lo referente a la ubicación original del refugio fue 109.4 ± 5.0% la rotación (25 respuestas de 8 ratones) y 109,1 ± 2.1%, rotación posterior (n = 8 respuestas de 8 ratones), p = 0.957 t-test entre pre y post-rotación, Figura 3A y B). Estos resultados demuestran que el proximal indicaciones visuales no se utilizan para guiar el vuelo hasta el refugio (pasos 8.3-8.5).
Cambiar la ubicación de la vivienda después de la obtención de una respuesta de escape exitoso (paso 6) resultó en ratones a la nueva ubicación de la vivienda poco después del cambio (tiempo promedio de visita el nuevo refugio = 33,1 s, gama = s 4-82). Posteriores ensayos secuenciales del sonido estímulo producción un aumento gradual en la probabilidad de escapar a la nueva ubicación (44.4% en el primer ensayo), alcanzando 100% en todos los animales por el quinto ensayo (n = 9 animales, figura 3). Estos experimentos de sondeo la dinámica de la actualización de la memoria de lugar de refugio para adaptarse a los cambios en el ambiente (paso 8.6).
Después de un período de aclimatación de 7 min al laberinto con ningún refugio (paso 7), la presentación de un estímulo visual tiempo de 5 s producido congelación comportamiento en lugar de escape (probabilidad de congelación = 95,2%, media de tiempo de congelación = 7,9 ± 2.7 s, n = 7 animales). Introducción posterior de un refugio en la misma sesión y la presentación del estímulo visual durante mucho tiempo produce escapar respuestas confiablemente, mostrando que ratones pueden cambiar flexiblemente estrategias defensivas dependiendo de la presencia de () refugio (figura 3D) pasos 8.1 y 8.7).

Figura 1: esquema de la arena de comportamiento. Dibujo de los componentes principales de la arena comportamiento visto desde la parte superior (derecha) y desde el lado (izquierda). Una gran parte de la plataforma se puede girar, incluyendo el refugio y el adjuntadas señales visuales. Círculos oscuros en la vista superior son los agujeros que el refugio se puede conectar, que son todos idénticos (se indica un lugar de refugio de ejemplo). La arena toda está encerrada en un armario de amortiguación de sonido. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: las respuestas de Escape son conductas intencionadas rápidos y precisos. (A) latencia al inicio de la fuga desde el inicio de la estimulación visual o auditiva. (B) la precisión de la navegación al refugio durante el escape, medida que el valor es 100% si el ratón se ejecuta directamente hasta el refugio y 10% menos por cada agujero del refugio (e.g., 80% si el agujero alcanzado primero fue dos agujeros de correcta). (C) desplazamiento del ratón durante escape conspiraron contra la distancia lineal entre la posición inicial del animal y la ubicación de destino, mostrando que el vuelo está cerca de linear. Caja y bigotes se muestran datos de 10-90 percentil y los restantes puntos de datos se muestran como círculos. Parcelas de color rojo son datos de estimulación auditiva (n = 36 respuestas de 15 animales) y el negro son de estimulación visual (n = 51 respuestas de 26 animales). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: ratones no utilizan proximal indicaciones visuales o señales relacionadas con la vivienda para guiar las respuestas de escape a. (A) parcela el desplazamiento como en la figura 2, para las respuestas pre- y post-rotación de la plataforma. (B) precisión de la navegación durante el escape en condiciones previas y post-rotación. Exactitud de la rotación posterior se mide en referencia a la ubicación inicial de la vivienda y muestra que los ratones navegación de nuevo a la ubicación original del refugio en su lugar la nueva posición. (C) fracción de respuestas de escape dirigidas hacia la nueva ubicación de refugio conspiraron contra número de estimulación. Después de la 5th presentación del estímulo, todos los animales escapan directamente hacia la nueva ubicación de la vivienda (n = 9 animales). (D) significa la velocidad a través de los animales en respuesta a la estimulación visual con refugio disponible (rojo) o ausente (azul), mostrando el escape y congelar las respuestas, respectivamente. Velocidad ha sido normalizada a los valores basales y alineado con tiempo de reacción (definida como sea el inicio de un vuelo o respuesta de congelación, línea discontinua). Las áreas sombreadas muestran SEM. (n = 7 animales). Para 2A y 2B, la caja y bigotes se muestran datos de 10-90 percentil y los restantes puntos de datos se muestran como círculos. Parcelas de color rojo son los datos de la estimulación auditiva antes de plataforma (n = 25 respuestas de 8 animales) y azul son las respuestas después de girar la plataforma (n = 8 respuestas de 8 animales). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
Los ensayos que se describen aquí son técnicamente fácil de realizar, y con la excepción de la plataforma de experimentos de rotación, se puede implementar fácilmente en un laberinto de Barnes estándar. Sin embargo, pueden surgir algunas complicaciones: por un lado, el ratón puede parecer tiene miedo de entrar en el refugio, que puede ser debido al refugio no está suficientemente limpia o seca. Por otro lado, el ratón puede permanecer dentro de la vivienda por períodos muy largos de tiempo. Es fundamental para estos experimentos que los animales nunca eliminan desde el refugio manualmente, ya que esto puede interrumpir el camino de integración así como la señal de socorro los animales y cambiar su respuesta a la amenaza. Se aconseja terminar el experimento después de 90 min y repetir el análisis después de 48 h si es necesario. Otra consideración práctica relevante es que cuando el ratón investiga el borde de la plataforma, no podrá detectar estímulos visuales si su cabeza está en ángulo sobre el borde, y por lo tanto recomendamos los estímulos en estos casos la retención. Además, ratones pueden saltar por la plataforma en raras ocasiones. Se aconseja terminar el experimento y repetición después de 48 h. ratones que activamente la plataforma una vez suelen hacerlo en el futuro y tengan que ser excluido del estudio. Finalmente, desde nuestra experiencia, la mayoría Ratones responderá a los estímulos visuales (34/36 ratones). Sin embargo, si los ratones no muestran ningún tipo de respuesta al estímulo visual (un susto, un vuelo o una respuesta de congelación), ellos deben ser excluidos del estudio.
Un punto importante a considerar es que el control de los estímulos sensoriales es normalmente un problema en los ensayos de memoria espacial, como a menudo es difícil eliminar todas las fuentes posibles de contaminación de señales. Nuestro montaje experimental reduce señales visuales externas que rodean el laberinto con una pared, y se coloca dentro de un gabinete de amortiguación de sonido para el aislamiento de sonido. Para minimizar las señales olfativas, recomendamos limpiar bien la instalación con etanol al 70% o ácido acético entre experimentos.
Nuestro análisis conductual se agrega a los métodos anteriores para estudiar navegación espacial26 utilizando conductas instintivas de defensa para sonda espacial memoria y centrarse en escenarios etológicas. Una diferencia crítica entre los procedimientos aquí descritos y el análisis estándar del BM es la falta de sesiones de entrenamiento. En nuestros análisis, el período de habituación asegura que el ratón visita el refugio de al menos una vez y a menudo muchas veces más, que anteriormente hemos mostrado suficiente memorizar la ubicación de vivienda2. Creemos que una razón importante por la alta tasa de éxito y precisión en la búsqueda de la vivienda a pesar de la falta de formación nunca pasivamente está desplazando al animal durante el experimento, haciendo integración vía una estrategia viable de la navegación. Sin embargo observamos que nuestro análisis se ocupa de memoria formado y evaluado durante una sola sesión, y que no hemos probado para la memoria a largo plazo de la localización del refugio, que suele ser el objetivo de los experimentos estándar de BM. Por último, utilizando estímulos innatamente aversivos discretos en lugar del ventilador utilizado o buzz para la duración de la sesión, nuestro análisis proporciona muy buen control experimental sobre dos conductas que podrían usar estrategias diferentes de navegación: búsqueda de alimento y dirigida por el refugio de escape. Creemos que el uso de estos ensayos en combinación con técnicas modernas de la neurociencia para la grabación y manipulación de la actividad de los nervios puede proporcionar penetraciones importantes en circuitos neuronales cómo calcular el comportamiento.
Divulgaciones
La mayoría de los datos incluidos en la sección de resultados es un subconjunto de los datos presentados en Vale et al 2017.
Agradecimientos
Este trabajo fue financiado por una beca de Wellcome Trust/Real Sociedad Henry Dale (098400/12/Z/Z), un Consejo de investigación médica (MRC) concesión de MC-UP-1201/1, Wellcome Trust y Gatsby Charitable Foundation SWC beca (T.B.), beca de doctorado de MRC (para D.E. y R.V) y una beca de doctorado de Fonds de Boehringer Ingelheim (para R.V). Agradecemos a Kostas Betsios para programar el software de adquisición de datos, la LMB mecánicos y talleres de electricidad para la construcción de la arena de la experimental.
Materiales
| Name | Company | Catalog Number | Comments |
| Infrared iluminators TV6700 | Abus | - | |
| DLP projector Infocus IN3126 | Infocus | 0001740992-00000001 | |
| Ultrasound speaker Pettersson L60 | Pettersson Elektronik | - | |
| Amplifier QTX PRO240 | QTX | - | |
| Soundcard Xonar D2 | Asus | 90-YAA021-1UAN00Z | |
| HP Z840 desktop | HP | F5G73AV | |
| 100 micron drafting film | Xerox | 3R98145 | |
| Near infrared camera: Basler acA1300-60gmNIR | Basler | 106202 | |
| National Instruments BNC-2110 | National instruments | 777643-01 | |
| LabVIEW 2015 64-bit | National instruments | - | |
| Custom made Barnes maze | MRC Laboratory of Molecular Biology mechanical workshop | - |
Referencias
- Eaton, R. C., Bombardieri, R. A., Meyer, D. L. The Mauthner-initiated startle response in teleost fish. J Exp Biol. 66 (1), 65-81 (1977).
- Vale, R., Evans, D. A., Branco, T. Rapid Spatial Learning Controls Instinctive Defensive Behavior in Mice. Curr Biol. 27 (9), 1342-1349 (2017).
- Blanchard, R. J., Flannelly, K. J., Blanchard, D. C. Defensive behavior of laboratory and wild Rattus norvegicus. J Comp Psychol. 100 (2), 101-107 (1986).
- Dill, L. M., Houtman, R. The influence of distance to refuge on flight initiation distance in the gray squirrel (Sciurus carolinensis). Canadian Journal of Zoology. 67 (1), 233-235 (1989).
- Blanchard, R. J., Blanchard, D. C., Rodgers, J., Weiss, S. M. The characterization and modelling of antipredator defensive behavior. Neurosci Biobehav Rev. 14 (4), 463-472 (1990).
- Fanselow, M. S., Lester, L. S. A functional behavioristic approach to aversively motivated behavior: Predatory imminence as a determinant of the topography of defensive behavior. Evolution and learning. , (1988).
- De Franceschi, G., Vivattanasarn, T., Saleem, A. B., Solomon, S. G. Vision Guides Selection of Freeze or Flight Defense Strategies in Mice. Curr Biol. 26 (16), 2150-2154 (2016).
- Bräcker, L. B., et al. Essential role of the mushroom body in context-dependent CO 2 avoidance in Drosophila. Current Biology. 23 (13), 1228-1234 (2013).
- Ghosh, D. D., et al. Neural architecture of hunger-dependent multisensory decision making in C. elegans. Neuron. 92 (5), 1049-1062 (2016).
- Bellman, K. L., Krasne, F. B. Adaptive complexity of interactions between feeding and escape in crayfish. Science. 221 (4612), 779-781 (1983).
- Rodgers, W., Melzack, R., Segal, J. "Tail flip response" in goldfish. Journal of comparative and physiological psychology. 56 (5), 917(1963).
- Dill, L. M. The escape response of the zebra danio (Brachydanio rerio) II. The effect of experience. Animal Behaviour. 22 (3), 723-730 (1974).
- Barnes, C. A. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 93 (1), 74-104 (1979).
- Pompl, P. N., Mullan, M. J., Bjugstad, K., Arendash, G. W. Adaptation of the circular platform spatial memory task for mice: use in detecting cognitive impairment in the APP(SW) transgenic mouse model for Alzheimer's disease. J Neurosci Methods. 87 (1), 87-95 (1999).
- Patil, S. S., Sunyer, B., Höger, H., Lubec, G. Evaluation of spatial memory of C57BL/6J and CD1 mice in the Barnes maze, the Multiple T-maze and in the Morris water maze. Behavioural Brain Research. 198 (1), 58-68 (2009).
- Sunyer, B., Patil, S., Höger, H., Lubec, G. Barnes maze, a useful task to assess spatial reference memory in the mice. Protocol Exchange. , (2007).
- Attar, A., et al. A shortened Barnes maze protocol reveals memory deficits at 4-months of age in the triple-transgenic mouse model of Alzheimer's disease. PLoS One. 8 (11), e80355(2013).
- Harrison, F. E., Reiserer, R. S., Tomarken, A. J., McDonald, M. P. Spatial and nonspatial escape strategies in the Barnes maze. Learn Mem. 13 (6), 809-819 (2006).
- Leary, T. P., Brown, R. E. The effects of apparatus design and test procedure on learning and memory performance of C57BL/6J mice on the Barnes maze. J Neurosci Methods. 203 (2), 315-324 (2012).
- Mittelstaedt, M. -L., Mittelstaedt, H. Homing by path integration in a mammal. Naturwissenschaften. 67 (11), 566-567 (1980).
- Stackman, R. W., Golob, E. J., Bassett, J. P., Taube, J. S. Passive transport disrupts directional path integration by rat head direction cells. J Neurophysiol. 90 (5), 2862-2874 (2003).
- Yilmaz, M., Meister, M. Rapid innate defensive responses of mice to looming visual stimuli. Curr Biol. 23 (20), 2011-2015 (2013).
- Mongeau, R., Miller, G. A., Chiang, E., Anderson, D. J. Neural correlates of competing fear behaviors evoked by an innately aversive stimulus. J Neurosci. 23 (9), 3855-3868 (2003).
- Hurst, J. L., West, R. S. Taming anxiety in laboratory mice. Nat Methods. 7 (10), 825-826 (2010).
- Paylor, R., Tracy, R., Wehner, J., Rudy, J. W. DBA/2 and C57BL/6 mice differ in contextual fear but not auditory fear conditioning. Behavioral neuroscience. 108 (4), 810(1994).
- Paul, C. M., Magda, G., Abel, S. Spatial memory: Theoretical basis and comparative review on experimental methods in rodents. Behav Brain Res. 203 (2), 151-164 (2009).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados