
Method Article
De Novo Génération de cellules souches somatiques par YAP/TAZ
Dans cet article
Résumé
Disponibilité de SCs somatiques est cruciale pour la médecine régénérative, modélisation de la maladie et de mieux comprendre les propriétés SC. Nous présentons ici des stratégies expérimentales de reprogrammer, in vitro, les cellules adultes différenciées dans les cellules correspondantes d’extensible tissu-spécifique souches/progénitrices par l’expression transitoire de l’activateur de transcriptional co seul YAP.
Résumé
Nous présentons ici des protocoles visant à isoler les cellules différenciées primaires et tourner en cellules souches/progénitrices (SCs) de la même lignée de l’expression transitoire du facteur de transcription YAP. Avec cette méthode, cellules luminales de (LD) différenciés de la glande mammaire de la souris sont converties en cellules qui présentent des propriétés moléculaires et fonctionnelles de SCs mammaire. YAP aussi tour à tour entièrement dissociés exocrine cellules pancréatiques dans le pancréas conduit comme progéniteurs. De même, à SCs endogènes, naturelles, induite par YAP souches comme cellules (« CJS ») peuvent être éventuellement étendus comme organoïde cultures à long terme in vitro, sans nécessité d’ectopique YAP/TAZ, comme CJS sont dotés d’un État autorenouvellement SC héréditaire.
La procédure de reprogrammation présentée ici offre la possibilité de générer et développez in vitro de cellules progénitrices de diverses sources de tissus à partir de cellules différenciées. L’expansion directe des cellules somatiques ex vivo a des implications pour la médecine régénérative, pour la compréhension des mécanismes d’initiation de la tumeur et, plus en général, pour la cellule et des études de biologie du développement.
Introduction
Spécifique aux tissus les cellules souches somatiques (SCs) sont essentiels pour renouvellement tissulaire et de la réparation après une blessure. La possibilité de facilement isoler et illimitée augmenter ex vivo somatique SCs représente un enjeu crucial pour thérapies régénératives potentiels, ainsi que pour les applications de SC en recherche fondamentale et de la modélisation de la maladie. Toutefois, les progrès dans cette direction, a été limitée par la difficulté de capturer l’état de SC de divers organes épithéliales in vitro. En effet, dans plusieurs tissus adultes résidents SCs peuvent existe pas, ou ne pas être facilement disponible, ou leur nombre et leur potentiel de régénération peuvent être érodés par les conditions de vieillissement ou une maladie. En 2016, nous avons commencé à combler cette lacune en signalant que l’expression d’un seul co-activateur transcriptionnel, YAP (protéine associée aux Oui) ou ses protéines apparentées TAZ (activateur de la transcription d’un motif PDZ), en cellules différenciées en phase terminale efficacement crée des populations de cellules fonctionnelles, extensible, non tumorigènes autologue qui sont sur le plan opérationnel et moléculairement indiscernables de leur correspondant du SCs tissu-spécifiques1. Une impulsion de subies YAP ou activité TAZ pour quelques jours est suffisante pour induire l’apparition du renouvellement automatique de SCs somatiques. Il s’agit d’un état stable qui n’est plus tributaire de l’expression du transgène continue, comme il peut être transmis par des générations de cellules sans davantage d’expression ectopique YAP/TAZ1. Le protocole présenté ici les détails la procédure utilisée pour générer des cellules souches/progénitrices épithéliales de novo de la glande mammaire et du pancréas, à partir de cellules différenciées de ces tissus. Cette procédure remplit une boîte noire dans l’arène de reprogrammation/transdifférenciation actuel. Principaux efforts dans ces directions ont en effet jusqu'à présent centrée sur la transition de la cellule à un état de cellules souches (iPSC) pluripotentes induites, suivie de la conversion de ces embryons et les cellules pluripotentes SCs en plus différencient. Toutefois, le CISP est tumorigènes une fois introduits dans les tissus adultes, soulevant la nécessité de développer des protocoles pour leur différenciation complète et efficace2. Toutefois, cette étape de différenciation, même lorsque cela est possible, est au prix de potentiels à long terme de repeuplement évolutivité, auto-organisation et orgue. Ce sont les qualités essentielles pour la régénération de l’orgue qui caractérisent en fait uniquement de SCs endogènes de tissu-spécifique et des SCs d’induite par YAP actuellement décrits (CJS). De même, transdifférenciation directe d’un type de cellule dans une autre à l’aide de cocktails de transcription divers facteurs génèrent également des cellules qui n’ont pas essentiel proliférative et stemness potentiels3différenciées.
La procédure décrite ici tire également parti de la technologie organoïde récemment introduite, par laquelle SCs endogènes peuvent être étendus et différencient ex vivo4. Induite par le YAP SCs peuvent générer SCs organoïde formant même dans des conditions biologiques ou maladie expérimentales, dans laquelle SCs endogènes ne sont pas présents. Nous tenons à noter que, à la différence avec d’autres procédures de reprogrammation, le type de la plasticité cellulaire communiquée par YAP peut correspondre à la seule forme de la réversion à un statut de SC-like qui se produit dans les tissus vivants. Réacquisition de SC-comme des traits a été associée à la réparation des tissus ou oncogènes activation5. Bien que dispensable pour l’homéostasie de plusieurs tissus adultes, YAP et/ou TAZ est absolument essentiel pour la régénération, la croissance tumorale et l’expansion de SCs somatiques in vitro1,6,7,8 ,9,10,11,12
Protocole
Toutes les procédures d’animaux ont été effectuées adhérant à nos lignes directrices et approuvée par OPBA et le ministère de la santé
1. génération de YAP-induced Mammary cellules souches (yMaSCs)
Remarque : Toutes les compositions de médias et de la solution pour la section 1 sont spécifiées dans le tableau 1.
- Isolement des populations de cellules mammaires primaires
- Préparer sous une hotte de culture cellulaire : solution hyaluronidase, moyen de dissociation, hémolytique solution, solution triage, Bistouris jetables, collagène, j’ai la solution de revêtement, Ca2 + solution de chélation, lavage moyen #1, lavage moyen #2, solution a, milieu de culture 2D mammaire, de la glace froide HBSS/PS.
- Pour une expérience typique, sacrifier 10 souris femelles (soit CD-1 de souche C57BL/6), âgés de 8 à 12 semaines par dislocation cervicale. Stériliser l’abdomen avec abondance de solution d’éthanol 70 % avant la dissection.
- Disséquer la glande mammaire par une incision en forme de Y le long de la peau abdominale et soigneusement séparant les glandes du péritoine en tirant doucement avec une pince Dumont. Placer les glandes disséquées dans un plat de cellules non adhésif avec de la glace 10 mL froid HBSS/PS (20 glandes pour chaque plat), faire attention à ne pas porter sur n’importe quel fragment de peau.
- Sous une hotte de culture tissulaire, laver chaque glande une fois dans 10 mL de frais HBSS/PS et placez-les dans un plat à adhésif non cellules vide (20 glandes pour chaque plat). Ne pas utiliser des plaques de culture de tissus, cellules auront tendance à s’en tenir à leur causant une perte importante de matériel.
- Finement hacher les glandes mammaires avec scalpels jusqu'à l’obtention d’un mélange homogène de fragments de 1 mm3 . Récupérer le tissu haché de chaque plat dans 10 mL de milieu de dissociation avec une pipette sérologique de 25 mL pour éviter l’encrassement et transférer la suspension dans un tube conique de 50 mL, pipetage au moins 5 fois pour désagréger les amas de tissu.
Remarque : Pour une digestion efficace, hachage correcte du tissu à l’étape 1.1.5 est crucial. - Incuber pendant 1 heure à 37 ° C sous agitation vigoureuse continue. Après 1 h, vérifier l’homogénat et prolonger l’incubation en 10 min si les touffes sont toujours présents. Tournez en bas les tissus digérés à 400 x g pendant 5 min à température ambiante et éliminer le surnageant. Resuspendre le culot de tissu dans 3 mL de solution hémolytique et incuber pendant 3 min sur la glace.
NOTE : Hémolyse est un traitement assez sévère, strict calendrier est donc crucial à cette étape. - Laver les cellules avec 10 mL de milieu de lavage #1, tournez en bas les tissus digérés à 400 g pendant 5 min et éliminer le surnageant. Resuspendre le culot de tissu dans 10 mL de lavage moyen # 2 et plaque en récipients de culture de tissu de 10 cm. Incuber les plats pendant 1 h à 37 ° C dans un incubateur de culture cellulaire.
Remarque : Cette étape permet l’élimination de la plupart des fibroblastes, qui doit adhérer à la boîte de Pétri. - Récupérer la suspension cellulaire de la vaisselle et les verser dans un tube conique de 50 mL. Tournant à 400 x g pendant 5 min et éliminer le surnageant. Laver le culot deux fois dans 10 mL de la Ca2 + solution de chélation par rotation vers le bas à 400 g pendant 5 min à chaque fois. Resuspendre le culot dans 5 mL de 0,25 % trypsine/EDTA et incuber pendant 5 min à 37 ° C.
- Ajouter 5 mL de solution a sur le dessus de la solution de trypsine et supplément à la DNase I 1 μg/mL. Pipette et descendre au moins 5 fois par un 1 mL-pointe à désagréger les amas de l’ADN et incuber pendant 10 min à 37 ° C, en secouant toutes les 3 min.
- Ajouter 10 mL de milieu de lavage #2 et le filtre dans un nouveau tube conique de 50 mL à travers un tamis de cellule 40 μm. Tournez en bas la suspension cellulaire à 400 x g pendant 5 min et éliminer le surnageant, en veillant à éliminer tout le liquide.
- Purification de cellules épithéliales mammaires par FACS
- Préparer le mélange d’anticorps en ajoutant 10 μL Lin (anticorps de lignée de souris cocktail), 12 μL anti-CD326 (Ep-CAM), à une concentration finale de 30 ng/mL, 10 μL anti-CD49f, à une concentration finale de 25 ng/mL, 10 μL anti-CD61, à une concentration finale de 10 ng/mL , 2,5 μL anti-CD29, à une concentration finale de 2,5 ng/mL.
Remarque : de cette étape à l’étape 1.3, toujours faire fonctionner dans l’obscurité, afin d’éviter le blanchiment des anticorps fluorescent étiquetés. Pour chaque pellet : - Garder un petit nombre de cellules (en plongeant une pointe de 100 μl dans le culot) dans une éprouvette. Remettre en suspension les cellules de 500 μL de solution dans un tube de FACS de tri et de garder sur la glace que l’échantillon non étiqueté pour la procédure de FACS.
- Resuspendre chaque culot dans 200 μl de milieu de lavage #1, ajouter 44,5 μl du mélange d’anticorps, Pipetter soigneusement et incuber pendant 30 minutes sur la glace dans l’obscurité. Diluer la suspension cellulaire dans 10 mL de milieu de lavage #1, tournez en bas à 400 x g pendant 5 min et éliminer le surnageant.
- Resuspendre le culot dans 2 mL de solution de tri et filtre à travers un bouchon-filtre FACS tubeand passez à FACS-séparation des populations cellulaires (comme dans la Figure 1 b). Avant d’effectuer le protocole de FACS multicolor, veillez à corriger l’effet d’entraînement possible de chaque fluorochrome dans chacune des autres. Pour ce faire, incuber chaque anticorps conjugué fluorophore séparément avec la suspension de cellules et de mesurer les valeurs spectrales chevauchement de fluorochromes et dans tous les détecteurs, par l’intermédiaire de contrôles de simple-couleur, afin de créer une matrice de compensation.
Remarque : Dans cette expérience, trieuse équipé 85 μm buse travaillait. Une préparation typique de 10 souris femelles produiront environ 800 000 cellules LD. Procéder à la filtration secondaire si touffes forment au cours de la procédure de FACS.
- Préparer le mélange d’anticorps en ajoutant 10 μL Lin (anticorps de lignée de souris cocktail), 12 μL anti-CD326 (Ep-CAM), à une concentration finale de 30 ng/mL, 10 μL anti-CD49f, à une concentration finale de 25 ng/mL, 10 μL anti-CD61, à une concentration finale de 10 ng/mL , 2,5 μL anti-CD29, à une concentration finale de 2,5 ng/mL.
- Ensemencement des cellules mammaires primaires de LD
- Au cours de la procédure de FACS recouvrir une plaque de culture de tissus bien multi avec collagène j’ai solution de revêtement. Incuber pendant 1 h à 37 ° C, 5 % de CO2 dans un incubateur de culture cellulaire. Enlever la solution de revêtement et laver avec du milieu de lavage #1 juste avant.
- Laver les cellules récupérées de la procédure de FACS avec 10 mL de solution de lavage #1, tournez en bas 400 g pendant 5 min et éliminer le surnageant. Resuspendre le culot dans le milieu de culture 2D mammaire (500 µL/puits) et plaque de collagène traité. Laisser les cellules dans un incubateur de culture cellulaire pendant 48 h permettre pour fixation des cellules appropriées et de diffusion.
Remarque : Pour un tri typique de 10 souris femelles, 6-8 puits d’une plaque bien multi 24 puits produira une densité cellulaire optimal (100 000 cellules/puits).
- Induction d’yMaSCs (YAP-induit des cellules souches mammaires)
NOTE : D’à partir de cette étape, toutes les procédures doivent être effectuées dans des conditions de BSL-2.- Préparer le milieu colonie mammaire. Fraîchement ajoute la matrice de la membrane basale au milieu, juste avant l’ensemencement de la cellule.
- Pour l’induction des cellules de tige mammaires induite par YAP (yMaSCs), transmettre les cellules primaires de LD par infection des gènes en mélangeant un volume de FUdeltaGW-rtTA surnageant viral, un volume de surnageant ICMSA-tetO-YAP (ou TAZ), avec deux volumes de mammaires sans sérum Milieu de culture 2D les concentrations de 2 x de suppléments, dans un volume total de 500 mL. Incuber les cellules avec des gènes surnageants pendant 48 h. Pour une préparation des gènes typique, veuillez consulter le protocole en ligne22.
- Après l’infection, laver les cellules adhérentes et traiter avec mammaire 2D culture additionné à 2 doxycycline µg/mL pour induire l’expression de gène exogène YAP (ou TAZ). Utiliser des cellules infectées par le vide, vecteurs exprimant EGFP ou YAPS94A ou les cellules infectées par des vecteurs inductibles de YAP (ou TAZ), mais laissées sans doxycycline comme témoins négatifs.
NOTE : Infection réussie peut être validée par qRT-PCR avec des amorces spécifiques pour homme YAP transgène, comme précédemment décrit1. - Après 7 jours d’induction avec la doxycycline, détacher les cellules adhérentes par incubation avec 0.05 % trypsine/EDTA (150 μl/puits) pendant 10 min à 37 ° C ; Arrêtez la trypsinisation en dilution 1:5 dans le milieu de lavage #2 (600 μL/puits) et compter les cellules. Remettre en suspension les cellules dans le milieu de la colonie mammaires (1 mL pour chaque puits), additionnés de 2 μg/mL doxycycline et la semence à une densité de clonogénique de 1 000 cellules/puits dans les plaques de fixation ultra basse 24 puits.
Remarque : Assurez-vous que le support de la colonie mammaire est glacé au moment de l’addition de matrice de membrane basale. La matrice de la membrane basale doit être conservée à-20 ° C à l’arrivée et décongelée lentement à 4 ° C durant la nuit ; une fois décongelé, il doit toujours être manipulé sur la glace, conformément aux directives du fabricant. - Une fois YAP-exprimant les cellules LD commencent à proliférer et se développer comme colonies MaSC-comme en suspension (colonies d’yMaSC) (14 jours après le semis), comte et traiter pour poursuivre l’analyse (Figure 1).
Remarque : Les cellules de contrôle négatif (comme dans le point 1.4.3) restera comme cellules individuelles. - Reconstituer la culture avec un milieu colonie mammaires frais toutes les 72 h durant les 14 jours de croissance des colonies yMaSC ; pour ce faire préparer une partie aliquote du milieu de la colonie mammaires sans membrane basale 5 % matrice, additionné de concentration 10 x de suppléments et ajouter 01:10 du volume total dans chaque puits (par ex. 100 μL dans 1 mL de milieu total), afin d’éviter une dilution excessive de la suspension de la matrice.
- Sub culture d’yMaSCs
- Récupérer les colonies primaires du milieu colonie mammaires, puis se dissocient et réamorcez.
Remarque : yMaSC colonies issues de cellules YAP-reprogrammé LD acquièrent la capacité d’auto-renouvellement et peuvent être avec succès sous – cultivées sans une administration de doxycycline (c'est-à-dire, indépendamment de l’expression des transgénique YAP/TAZ) plus loin. - Préparer, sous une hotte de culture cellulaire, milieu de la colonie mammaires comme au point 1.4.1 et mammaires organoïde milieu.
- Collecter chaque échantillon et incuber dans l’excès de volume (10:1) de la glace froide HBSS pendant 1 h sur la glace, afin de solubiliser la matrice de la membrane basale. Laver les colonies 3 x par centrifugation à 180 x g pendant 5 min et remettre dans la glace froide HBSS. Incuber les colonies à 0.05 % trypsine/EDTA pendant 10 min à 37 ° C pour obtenir une suspension monocellulaire. Colonies de pipette et descendre de 10 x avec une pointe de p1000 pour assurer une dissociation complète au niveau de la cellule unique.
- Comte et réensemencer les cellules dans le milieu de la colonie mammaires (1 mL pour chaque puits) sans la doxycycline à une densité de clonogénique de 1 000 cellules/puits dans les plaques de fixation ultra basse 24 puits. Répétez ce passage procédure tous les 10-14 jours pour évaluer l’auto-renouvellement.
- Avant le troisième passage, passage yMaSC colonies en conditions de culture organoïde, afin d’améliorer yMaSC expansion et pour permettre la formation de mini-glandes, qui s’organiser dans un épithélium double étroitement réminiscent de la glande en vivo Organisation histologique.
- Récupérez les colonies depuis le milieu de la colonie mammaires comme au point 1.5.3 à 1.5.4. Colonies de remettre au facteur de croissance de 100 % réduit la matrice de la membrane basale, considérant à replate un maximum de 20-25 colonies pour chaque puits d’une plaque de fixation ultra basse 24 puits dans 150 µL de la matrice.
- Incuber les boîtes dans un incubateur de culture cellulaire pendant 40 min à 37 ° C et laissez la matrice de la membrane basale se solidifier et ensuite superposer les gels avec 500 μl de milieu organoïde mammaire.
- Après quelques jours, vérifier pour la formation de colonies pour former en herbe organoïdes (Figure 1E).
- Après 10-14 jours, passage ou processus l’organoïdes pour une analyse ultérieure.
- Au passage organoïdes cultures, récupérer organoïdes en recueillant chaque échantillon et d’incubation dans un volume excessif (10:1) de la glace froide HBSS pendant 1 h sur la glace, afin de solubiliser la matrice de la membrane basale. Laver organoïdes 3 x par rotation vers le bas à 180 x g pendant 5 min et remettre en suspension dans la glace froide HBSS.
- Incuber organoïdes à 0.05 % trypsine/EDTA pendant 10 min à 37 ° C pour obtenir une suspension monocellulaire. Pipette organoïdes haut en bas 10 x avec une pointe de p1000 pour assurer une dissociation complète au niveau de la cellule unique.
- Réamorcez sous forme de suspension monocellulaire dans une goutte de matrice réduite membrane basale de facteur de croissance de 100 % (150 μL de chaque puits d’une plaque de fixation ultra basse 24 puits). Laissez la matrice de la membrane basale forment un gel en incubant 40 min à 37 ° C dans un incubateur de culture cellulaire et superposer les gels avec 500 μl de milieu organoïde mammaire.
Remarque : yMaSC organoïdes peut être cryoconservés en récupérant de la culture de matrice de membrane basale de 100 % comme au point 1.5.13, évitant la trypsinisation. Stocker dans le mammaires organoïde additionné de 10 % DMSO. - Congeler rapidement l’organoïdes yMaSC à-80 ° C et ensuite conservés dans l’azote liquide.
- Récupérer les colonies primaires du milieu colonie mammaires, puis se dissocient et réamorcez.
2. génération d’yDucts
Remarque : Toutes les compositions de médias et de la solution au chapitre 2 sont indiquées au tableau 2.
- Isolement des acini pancréatiques primaires
- Placer les ciseaux et pinces de dissection dans 70 % EtOH et préparer sous un hotte acineuses culture milieu de culture cellulaire, 15 mL pour chaque souris ; support de récupération acineuses, 60 mL pour chaque souris ; PBS/PS ; collagénase de solution mère j’ai ; collagénase j’ai solution A, 15 mL pour chaque souris ; neutralisé queue-de-rat collagène j’ai solution. Neutraliser le collagène de queue de rat I à pH = 7, en réglant d’abord avec NaOH 0,1 N à l’acide acétique dans laquelle le collagène se dissout, puis avec 10N HCl. diluer à 2,5 mg/mL de tampon en PBS/PS garder le collagène de queue de Rat I et tous les réactifs sur la glace pour le neutraliser. Sacrifier 6 à 9 semaines-vieille souris du génotype adéquat.
- Placez chaque souris sur son dos et laver l’abdomen avec une solution d’éthanol à 70 %. Faire une incision longitudinale le long de la paroi abdominale. Localiser et disséquer le pancréas (en utilisant la rate comme guide) et le placer dans un plat adhésif non cellules de 10 cm dans la glace de 10 mL PBS/PS-transfert froid les plats immédiatement sous une hotte de culture de cellules. Travailler toujours de partir de cette étape, sous une hotte de culture cellulaire.
- Transférer chaque pancréas dans une nouvelle cellule non adhésif plat précédemment rempli de 7 mL de collagénase j’ai solution A.
- Rapidement, émincer chaque pancréas avec une paire de Bistouris jetables, pour obtenir une suspension homogène de tissu de fragments de3 environ 1 mm.
Remarque : Il est important de noter que cette procédure devrait prendre pas plus de 2 min pour la viabilité cellulaire optimal. - Incuber le plat pour la digestion de collagénase à 37 ° C, 5 % de CO2 dans un incubateur de culture cellulaire pendant 10 min, secouant toutes les 3 min pour assurer la digestion de tissu homogène.
- Récupérer les tissus digérés dans un tube conique de 50 mL (un pour chaque pancréas), laver le plat avec 10 mL de milieu de lavage acineuses et placez-le dans le même tube conique de 50 mL, pipetage digéré tissu haut et bas pas plus de 3 x.
- Tournez en bas les tissus digérés pendant 5 min à 100 g à 18 ° C et éliminer le surnageant.
Remarque : Tournez en bas les cellules à 18 ° C pour réduire l’activité de la collagénase au cours de cette étape. - Resuspendre le tissu pellet dans 7 mL de collagénase j’ai solution A et verser cette solution dans une autre boite de colle de cellules non 10 cm.
- Incuber le plat pour une deuxième série de digestion de collagénase à 37 ° C pendant 10 min comme au point 2.1.5, secouant toutes les 3 min pour assurer la digestion de tissu homogène. Pendant ce temps, préparer un tube conique propre 50 mL pour chaque pancréas surmonté d’une crépine de cellule 100 μm.
- Récupérer les tissus digérés et passer à travers la crépine de cellule de 100 μm par macération des tissus avec un piston de seringue stérile de 10 mL (veillez à enfoncer avec soin le tissu, évitant les forces de cisaillement tangent à la surface du tamis). Laver le plat avec 10 mL de milieu de lavage acineuses et traverser ce 10 mL de la crépine de cellule μm 100 même.
- Tournez en bas les tissus digérés pendant 5 min à 100 g à 18 ° C et éliminer le surnageant.
- Récupérer la pastille de tissu avec 10 mL de milieu de lavage acineuses. Transférer la solution de cellules dans un tube conique de 50 mL contenant supplémentaire 10 mL de milieu de lavage acineuses frais déjà, en évitant pipetage excessive pour la remise en suspension de la pastille.
- Tournez en bas les tissus digérés pendant 5 min à 100 g à 18 ° C et éliminer le surnageant.
- Soigneusement Resuspendre les tissus digérés dans 6 mL de milieu de récupération acineuses et distribuez-le dans les 2 puits de 6 puits multi bien vitroplants plaque, 3 mL chacun. Sous un stéréomicroscope soigneusement évaluer la qualité de l’isolement acineuse, qui apparaissent sous forme d’une suspension homogène de clusters acineuses, avec une faible proportion de cellules uniques (voir la Figure 2 b) ; supprimer n’importe quel tissu gros amas éventuellement présent (généralement visible à le œil nu), par pipetage eux hors de la solution.
- Ensemencement des acini pancréatiques primaires
- Incuber les clusters acineuses digérées à 37 ° C dans un incubateur de culture cellulaire pendant 2 h, pour permettre la récupération de la cellule.
- Dans cette couche de récupération cellule multi 48 puits puits avec 100 μL de collagène neutralisée rat queue j’ai et incuber pendant 1 heure à 37 ° C dans un incubateur de culture cellulaire pour permettre un coussin d’hydrogel pour former.
- Après 2 h de récupération de la cellule, recueillir la suspension de cellules acineuses dans un tube à fond conique, tournez vers le bas pendant 5 min à 100 g à 18 ° C et éliminer le surnageant.
- Remettre en suspension les acini dans le volume approprié de milieu de culture acineuse (150 μL à chaque puits d’une plaque de culture de tissus bien 48). Semences chaque dissociés du pancréas en 16 puits afin d’obtenir une densité optimale (100-120 acineuses grappes/puits).
- Diluer cette suspension acinaire avec un volume égal de collagène de queue de rat neutralisée j’ai solution, garder les tubes sur la glace. Mélanger soigneusement et rapidement des graines la suspension cellulaire sur le coussin de collagène décrit à la section 2.2.2 (300 μl à chaque puits d’une plaque de culture de tissus bien multi bien 48).
- Incuber 1 h à 37 ° C dans un incubateur de culture cellulaire pour permettre un hydrogel pour former.
- Induction d’organoïdes pancréatique
- Superposition des hydrogels de collagène avec 500 μL d’acineuses Culture additionné à 2 doxycycline μg/ml pour l’induction de YAP dépendant du pancréas organoïdes de R26-rtTAM2 ; TetO-YAPS127A souris ; contrôles négatifs sont fournis par wt cellules cultivées dans les mêmes conditions ou R26-rtTAM2 ; TetO-YAPS127A cellules cultivées en absence de doxycycline.
- Cellules acineuses de culture dans le milieu de Culture acineuses additionné de 2 doxycycline μg/mL pendant 5 à 7 jours rafraîchissant de milieu de culture (300 μL/puits) toutes les 48 h et après formation organoïde par des changements morphologiques vers kyste formant des structures ressemblant à des canalaire ( Figure 2). Une fois que les organoids se forment, cellules peuvent être repiquées dans des conditions de culture organoïde pancréatique ou récoltés pour approfondir les analyses (par exemple: extraction de l’ARN, immunofluorescence).
- Sub culture d’organoïdes pancréatique
- Afin d’évaluer leur capacité d’autorenouvellement, par clonage passage organoïdes pancréatiques induite par YAP (yDucts) dans la membrane de sous-sol en trois dimensions matrice hydrogels (conditions de culture organoïde pancréatique) indépendamment l’apport exogène de YAP/TAZ (i.e. indépendamment de l’administration doxycycline).
- Préparer la trypsine 0,05%/EDTA ; matrice de facteur de croissance de 100 % de réduction membrane basale ; Pancréatique organoïde Medium et collagénase j’ai solution B.
- Préparer un 15 mL conique tube avec 4 mL de collagénase, j’ai la solution B pour chaque puits être repiquées.
- Jeter le milieu de culture, soigneusement extrait des puits hydrogels en aspirant doucement et transférez-les sur les tubes coniques.
- Incuber les tubes à 37 ° C pendant 30 min, avec une agitation vigoureuse continu pour permettre une digestion complète de la matrice de collagène (cocher toutes les 10 min jusqu'à ce que l’hydrogel est complètement solubilisé). Tournez en bas des cellules récupérées à 750 g pendant 2 min et éliminer le surnageant.
- Incuber les cellules récupérées dans 1 mL de trypsine 0,05%/EDTA pendant 10 min à 37 ° C pour obtenir une suspension monocellulaire. Diluer la trypsine avec 9 mL de PBS 1 x, tournez en bas à 750 g pendant 2 min et éliminer le surnageant.
- Resuspendre le culot dans la matrice de glace froide facteur de croissance réduite de membrane basale et semences dans des plaques de fixation ultra faible (généralement une baisse de 150 μL dans un puits d’une plaque 24 puits).
- Laisser la membrane basale matrice hydrogel se solidifient en incubant les plaques dans un incubateur de culture cellulaire 40 min à 37 ° C et ensuite overlay avec pancréatique organoïde moyen (500 μl à chaque puits). yDucts se développera comme kyste-comme organoïdes en 7-10 jours (Figure 2D).
- Pour le passage plus organoïdes peut être retiré de la matrice de la membrane basale par incubation dans le froid glace PBS 1 x pendant 30 min, ensuite laver 3 fois par rotation à 180 x g pendant 5 min et remise en suspension dans du PBS froid glace 1 x afin d’éviter le report de la matrice. Organoïdes sont alors dissociées avec la trypsine 0.05 % pendant 10 min obtenir une suspension de cellules individuelles et redéfinies dans la matrice de la membrane de sous-sol frais et puis recouvert le pancréas organoïde moyen (comme dans 2.4.7-2.4.8).
- yDuct organoïdes peut être cryoconservés dans l’azote liquide en récupérant de membrane basale 100 % culture de matrice comme à l’étape 2.4.9, évitant la trypsinisation, et stocker dans un milieu organoïde pancréatique additionné de 10 % DMSO.
- yDuct organoïdes sont rapidement congelés à-80 ° c et ensuite conservés dans l’azote liquide.
Résultats
Génération d’yMaSCs
Un aperçu de la stratégie expérimentale pour reprogrammer les cellules mammaires primaires de LD par expression transitoire de YAP est présenté dans la Figure 1 a. Principales cellules épithéliales mammaires de la LD sont purifiés par fluorescence-lancée de cellules tri13. Figure 1 b représente une procédure de tri typique pour obtenir trois sous-populations distinctes : cellules basales (EpCAMfaibleCD49fhauteCD61–), les cellules progénitrices luminale (LP) (EpCAMhauteCD49ffaibleCD61+ ) et les cellules de LD (EpCAMhauteCD49ffaibleCD61–). Attention blocage des trois sous-populations est essentiel pour isoler une pure préparation de cellules LD, qui se distinguent entièrement et complètement croissance arrêtée lorsqu’ensemencées en formant des conditions (voir la Figure 1, panneau latéral gauche) des colonies de la glande mammaire. À l’inverse, lorsque amené à exprimer YAP exogène, cellules de LD commencent prolifèrent pour former des colonies épithéliales denses facilement reconnaissables dans les cultures en suspension matrice membrane basale 5 % (Figure 1). L’efficacité de la reprogrammation, attesté autour de 3 % pour une expérience, peut être marqué en comptant le nombre de colonies sur le nombre de cellules individuelles initialement ensemencé dans la membrane basale des cultures en suspension matrice (Figure 1). Les cellules luminales reprogrammés (yMaSCs) peuvent alors être passage dans 100 % membrane basale matrice organoïde conditions de culture (voir Figure 1 a), auto-organisés dans des structures ressemblant à des organoïde complexes qui se développent autour des lumières multiples et afficher la remarquable capacité d’autorenouvellement même en absence de doxycycline (c'est-à-dire en l’absence d’expression de YAP transgénique) (Figure 1E). Sur le plan histologique, dérivé d’yMaSC organoïdes affichent une couche basale (K14 positive), face à la matrice de la membrane basale reconstituée ECM et une couche luminale (K8 positive), face à des cavités lumen-comme dans l’organoïde (Figure 1F). Cette architecture est indiscernable de celle d’organoïdes formé par MaSCs natif (Figure 1F).
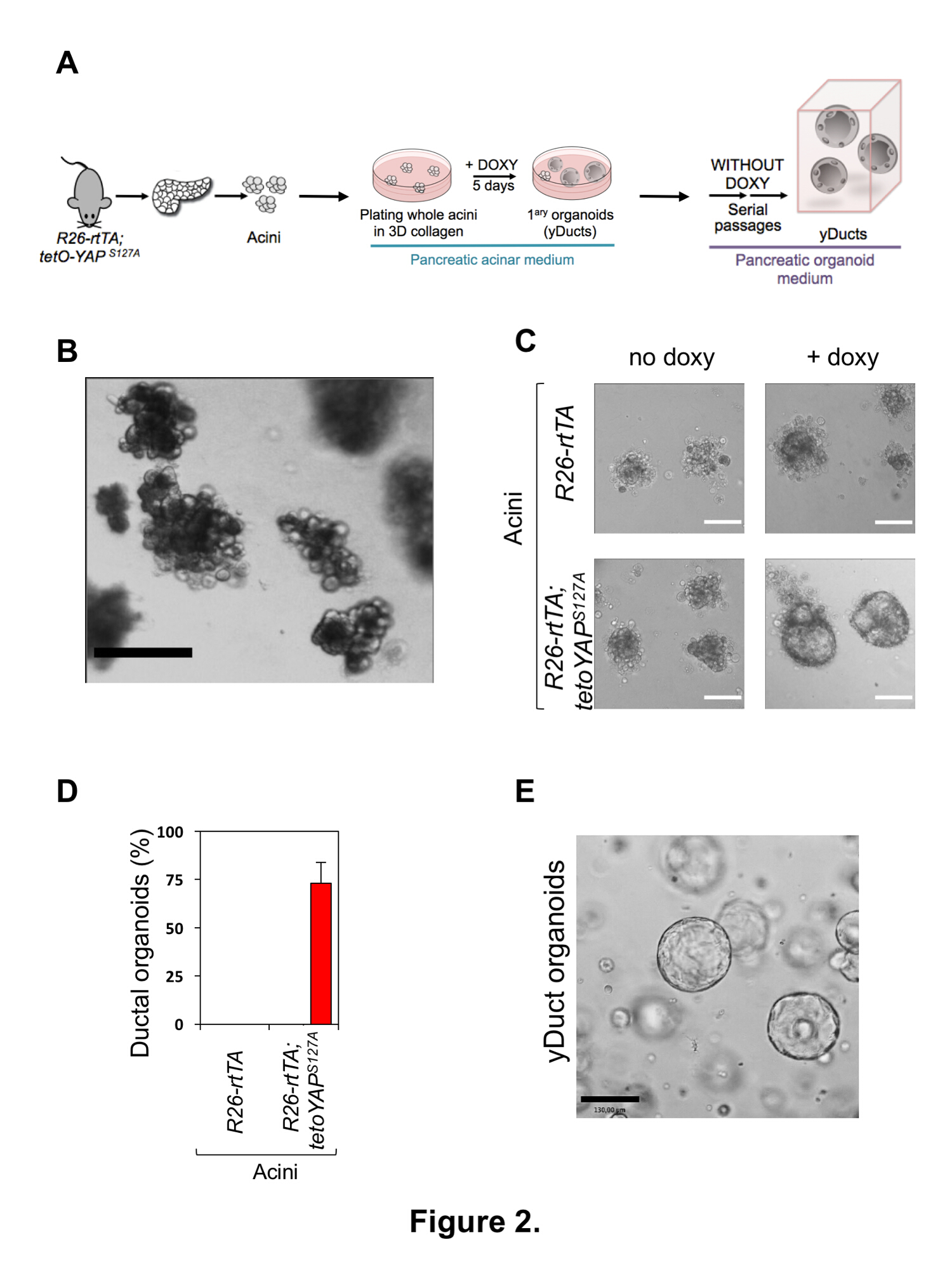
Génération d’yDucts
Un aperçu de la stratégie expérimentale pour reprogrammer les acini pancréatiques primaires par l’expression transitoire de YAP est présenté dans la Figure 2 a. Toute acineuses clusters sont isolés de la masse du tissu pancréatique par une combinaison de dissociation douce et l’exclusion de taille par filtration. Une préparation typique est présentée dans la Figure 2 b. Après l’isolement, les grappes de cellules acineuses doivent apparaître sous forme de suspension exocrine acineuses unités de taille homogène, sans contamination par endocrinien îlots de Langerhans ou fragments de l’arborescence ductal pancréatique et dissociation minime aux cellules individuelles. Contamination par des îlots endocriniens ou fragments canalaires est une indication de filtration sélective déficiente (étape 2.1.10), probablement à cause de traitement rigoureux ; dissociation indésirable de clusters acineuses à monocellules peut-être dû à traitement de collagénase excessive ou non l’activité des enzymes protéolytiques libéré par le tissu, qui peut être freiné par un traitement additionnel SBTI.
Une expérience typique de reprogrammation acineuse est présentée dans la Figure 2; dans 5-7 jours de culture en collagène 3D-je me suis basé hydrogel en présence de doxycycline, les acini pancréatiques dérivés R26-rtTAM2/tetO-YAPS127A souris facilement se transformer en amas semblables conduit (que nous avons appelée yDucts), composée par un mince monocouche de cellules épithéliales qui prolifèrent autour d’une cavité centrale en pleine expansion. La reprogrammation de l’efficacité, qui représente environ 70 % pour une expérience typique, peut être facilement mesurée en marquant le nombre de clusters conduit comme sur le nombre total de graines acini (Figure 2D). Cellules de contrôle négatif, c'est-à-dire R26-rtTAM2 / + cellules ou cellules R26-rtTAM2/tetO-YAPS127A gauche sans doxycycline, invariablement restent comme post-mitotiques clusters acineuses dans ces conditions de culture, comme indiquées précédemment1 ,14,15. YDucts reprogrammé peut ensuite être repiquées au niveau unicellulaire dans de conditions culture axée sur le Matrigel organoïde16 (voir Figure 2 a), affichage remarquable capacité d’autorenouvellement même en absence de doxycycline (c.-à-d. en l’absence d’expression de YAP transgénique) (Figure 2E).

Figure 1 : Isolement de LD mammaire primaire des cellules et l’induction de mammaire de cellules souches. (A) représentation schématique de la procédure expérimentale adopte pour reprogrammer les cellules mammaires primaires de LD. (B) représentant FACS-parcelles illustrant une procédure de tri typique pour purifier les cellules de la LD. i) cellules dissociées sont bloquées après dispersion vers l’avant et latérale pour des cellules vivantes (P1 ; bleu) ; II) population P1 est alors plus dépendants selon son profil de Lin : la sous-population de cellules de la lignée négatives (P2 ; gris) est sélectionnée, à l’exclusion des cellules hématopoïétiques lignée positives ; III) population P2 est alors séparée en un EpCAMhaute (P3 ; jaune + vert) et un sous-populationsfaible (P6 ; rouge) EpCAM ; IV) P3 et P6 sont alors davantage dépendants selon leur profil CD61/CD49f en trois sous-populations : EpCAMfaibleCD49fhauteCD61 cellules basales de– (P7 ; rouge), EpCAMhauteCD49ffaibleCD61+ LP cellules (P8 ; jaune) et EpCAMhauteCD49ffaibleCD61 cellules de LD– (P9 ; vert). (C) Images illustrent la capacité des cellules de LD, infectées par les constructions indiquées, pour former des colonies mammaires 15 jours après l’ensemencement dans le milieu de la colonie mammaire. Seulement tour de cellules exprimant le YAP en formant des colonies cellules, tandis que les cellules contrôle négatif (EGFP-infectés) demeurent comme arrêté à la croissance des cellules individuelles. Echelle = 50 μm. (D) la Quantification de la colonie formant la capacité des cellules indiquées, comme en (C). Les données sont présentées comme moyenne + s.d. et sont représentatifs des cinq expériences indépendantes, chacune avec six réplicats techniques. (E) l’image représentative de YAP-reprogrammé cellules souches comme cellules mammaires (yMaSCs) après 12 jours en conditions de culture organoïde dans trois dimensions frais 100 % membrane basale matrice hydrogel en l’absence de la Doxycycline. Echelle = 100 μm. (F) images représentatives immunofluorescence pour le marqueur basal K14 (vert) et le marqueur luminal K8 (rouge) d’organoïdes a dérivé les cellules indiquées, après 12 jours en conditions de culture organoïde. Echelle = 10 μm. Cette figure est reproduite de Panciera et coll., 20161. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 : Isolement des cellules acineuses du pancréas primaires et induction des cellules souches pancréatiques. (A) représentation schématique de la procédure expérimentale adoptée pour reprogrammer des cellules acineuses exocrines pancréatiques primaires. (B) l’image représentative des acini pancréatiques primaires juste après la procédure d’isolement (étape 2.1.14). La préparation acineuse doit apparaître comme une suspension homogène de grappes acineuses, avec une présence minimale des cellules individuelles. Echelle = 400 μm. (C) des images représentatives des acini pancréatiques primaires issus de la R26-rtTAM2 (panneaux supérieurs)ou R26-rtTAM2 ; tetO-YAPS127A (baisse de panneaux) souris et cultivées en 3D collagène I-base d’hydrogel pour 5 jours avec ou sans Doxycycline (doxy), comme indiqué. Seulement exprimant le YAP primaire acini convertit en cellules cultivées comme les kystes organoïde après addition de la Doxycycline. Barreaux de l’échelle = 70 μm. (D) la Quantification de la capacité des acini pancréatiques pour former organoïdes canalaire à la surexpression de YAP transgénique comme en (C). Les données sont présentées en moyenne + s.d. et sont représentatifs des cinq expériences indépendantes, effectuées avec quatre répétitions techniques. Image de représentant (E) des cellules de type canalaire YAP-reprogramed (yDucts) après trois passages dans trois dimensions frais 100 % membrane basale matrice hydrogel en l’absence de Doxycycline. Echelle = 130 μm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
| Isolement des cellules mammaires primaires | |
| Ca2 + solution de chélation | Conserver à 4 ° C |
| EDTA | 0,02 % p/V |
| PBS | |
| Collagène I solution de revêtement | |
| Acide acétique 0.02N, pH 3,23 | |
| Collagène de queue de rat (revêtement) | 01:50 |
| Solution a | Conserver à 4 ° C |
| A | 5 mg/ml |
| PBS | |
| Moyen de dissociation | |
| DMEM:F12 | |
| Solution mère de l’hyaluronidase | 400 U/mL |
| Stylo/Strep | 1 x |
| Collagénase de solution mère j’ai | 600 U/mL |
| Solution hémolytique | Conserver à 4 ° C |
| Solution de NH4Cl | 1 pièces |
| TrisBase 20,6 g/L | 9 pièces |
| ajuster le pH à 7,2 | |
| HBSS/PS | Conserver à 4 ° C |
| HBSS | |
| Stylo/Strep | x 2 |
| Solution mère de l’hyaluronidase | Filtre 0,2 µm, conserver à 4 ° C |
| Hyaluronidase des testicules bovins (poudre) | 2 000 U/mL |
| 1 7,3 M de tampon phosphate de sodium | |
| Solution de NH4Cl | Stocker à T. amb. |
| H2O | |
| NH4Cl | 7,1 g/L |
| ajuster le pH à 7,65 | |
| Solution de tri | Filtre 0,2 µm, conserver à 4 ° C |
| BSA | 0,1 % |
| EDTA | 1 mM |
| PH HEPES 7 | 25 mM |
| PBS | |
| Laver le moyen #1 | |
| DMEM/F12 | |
| Stylo/Strep | 1 x |
| Laver le moyen #2 | |
| DMEM/F12 | |
| FBS | 5 % |
| Stylo/Strep | 1 x |
| Milieu de culture 2D mammaire | |
| DMEM/F12 | |
| FBS | 2 % |
| héparine | 4 mg/mL |
| L-Glutamine | 1 x |
| bFGF murin | 10 ng/mL |
| EGF murin | 10 ng/mL |
| Stylo/Strep | 1 x |
| Induction et Passaging d’yMaSCs | |
| Moyen de colonie mammaire | |
| DMEM:F12 | |
| FBS | 5 % |
| héparine | 4 µg/mL |
| L-Glutamine | 1 x |
| Matrigel (ajouter immédiatement avant le semis) | 5 % |
| bFGF murin | 20 ng/mL |
| EGF murin | 10 ng/mL |
| Stylo/Strep | 1 x |
| Mammaires organoïde Medium | |
| DMEM:F12 avancée | |
| B27 | 1 x |
| GlutaMax | 1 x |
| héparine | 4 µg/mL |
| HEPES | 1 x |
| Caboche humaine | 100 ng/mL |
| bEGF murin | 20 ng/mL |
| EGF murin | 50 ng/mL |
| R-Spondin 1 | 1 µg/mL |
Tableau 1 : génération d’yMaSCs. Composition de tous les différents milieux de culture et les solutions requises pour l’isolement de cellules primaires de LD mammaires et induction d’yMaSCs (section 1)
| Isolement des acini pancréatiques primaires | |
| Milieu de Culture acineuses | |
| BPE | 50 µg/mL |
| BSA | 0,1 % |
| Dexaméthasone | 1 µg/mL |
| FBS | 0,1 % |
| ITS-X | 1 x |
| Stylo/Strep | 1 x |
| SBTI | 0,2 mg/mL |
| Moyen de Waymouth | |
| Support de lavage acineuses | |
| BSA | 0,1 % |
| Stylo/Strep | 1 x |
| Milieu RPMI | |
| SBTI | 0,2 mg/mL |
| Moyen de récupération acineuses | |
| Milieu de Culture acineuses | |
| FBS | 30 % |
| Collagénase j’ai solution A | |
| Support de lavage acineuses | |
| Collagénase de solution mère j’ai | 360 U/mL |
| PBS/PS | Conserver à 4 ° C |
| PBS (tampon Phosphate salin) | |
| Stylo/Strep | 1 x |
| Collagénase de solution mère j’ai | Conserver à-20 ° C |
| Collagénase, type I (poudre) | 6 000 U/mL |
| PBS | |
| Passage des organoïdes pancréatique | |
| Collagénase j’ai solution B | |
| PBS 1 x | |
| Collagénase de solution mère j’ai | 240 U/mL |
| Pancréatique organoïde Medium | |
| DMEM avancé/F12 | |
| B27 | 1 x |
| gastrine | 10 nM |
| FGF10 humaine | 100 ng/mL |
| Caboche humaine | 100 ng/mL |
| EGF murin | 50 ng/ml |
| N-acétylcystéine | 1,25 mM |
| Nicotinamide | 10 mM |
| Stylo/Strep | 1 x |
| R-Spondin 1 | 1 mg/mL |
| SBTI | 0,2 mg/mL |
Tableau 2 : génération d’yDucts. Composition de tous les différents milieux de culture et les solutions requises pour l’isolement des cellules acineuses primaires et l’induction et le passage des yDucts (section 2)
Discussion
Ici, nous présentons des protocoles pour reprogrammer les ex vivo terminalement différenciées des cellules épithéliales des tissus différents dans leurs cellules progénitrices spécifiques des tissus correspondantes (ou CJS) par une expression transitoire de YAP, tel que rapporté précédemment1. Nous avons détaillé les deux procédures : une permettant la reprogrammation de cellules FACS purifié par le biais de vecteurs LENTIVIRAUX et une seconde qui évite l’infection virale et tire profit de l’expression de YAP transgénique. Chaque protocole présente une stratégie efficace pour isoler et cellules différenciées principal de culture et d’une stratégie pour forcer exogène YAP l’expression des gènes dans les cellules différenciées, générant de novo spécifique aux tissus extensibles les cellules souches somatiques (voir schémas dans les Figures 1 a et 2 a).
Nous avons démontré que les stratégies d’isolement présentés ici effectivement isoler une population pure de cellules différenciées, comme en témoigne le fait que nous avons jamais détecté toute excroissance provenant d’échantillons de contrôle négatif (Figures 1 et 2 C).
Les vecteurs LENTIVIRAUX utilisés dans cette étude pour la reprogrammation des cellules mammaires primaires de LD sont doxycycline inductible, offrant la possibilité d’un contrôle strict de l’expression du transgène ; Cela permet de mettre en marche et arrêt YAP exogène expression à volonté. Une attention particulière doit être placée en évitant l’utilisation d’un titre viral excessive, comme cela peut être néfaste en termes d’efficacité de reprogrammation. Dans le cas des cellules acineuses primaires, nous sommes passés à une approche entièrement transgénique afin d’obtenir une reprogrammation de YAP-dépendante avec les manipulations minimes. Cette dernière stratégie est aussi particulièrement indiquée pour les acini pancréatiques primaires, comme groupes acineuses isolés sont guère propices à l’infection des gènes et très fragile. La stratégie transgénique employée offre l’avantage même des vecteurs LENTIVIRAUX doxycycline-dépendante pour le contrôle strict de l’expression génique. En outre, la stratégie transgénique exploitée avec les acini pancréatiques primaires porte l’avantage supplémentaire d’un rendement beaucoup plus élevé reprogrammation par rapport à l’origine virale de reprogrammation des cellules mammaires de LD. Au-delà de la plasticité intrinsèque différente associée aux cellules provenant de tissus différents, le taux supérieur de reprogrammation du pancréas pourrait provenir de l’efficacité supérieure d’expression liée à l’expression de YAP uniforme et autonome dans tous les cellules explantées. Notamment, nous avons démontré que YAP exogène n’est plus nécessaire après la génération du CJS (colonies d’yMaSC et yDucts), sans affecter leur capacité d’autorenouvellement. C’est parce que le CJS réactivation endogène YAP/TAZ et utilisent pour autorenouvellement lorsque YAP exogène est éteint1.
Nous avons validé la notion que les CJS sortent en effet des cellules différenciées en contrôlant la cellule d’origine de nos expériences de reprogrammation génétique lineage-traçage validations1.
Caractérisation desde CJS montre qu’induite par YAP reprogrammation génère normal somatique SCs1 comme je l’ai) niveau transcriptomique, CJS affichent massive chevauche SCs natifs ; II) CJS potentiel de différenciation d’affichage et peuvent générer une descendance multilignée toujours limitée à l’identité de leur tissu d’origine ; III) CJS sont non transformées et non-tumorigènes quand transplanté en vivo.
Ici on décrit aussi les procédures visant à maintenir et à développer dans la culture fois yMaSCs et yDucts comme organoïdes incorporé dans la membrane basale 100 % matrice hydrogels. Ces conditions permettent l’auto-organisation du CJS en organoïdes en trois dimensions qui assurent le maintien des propriétés stemness à long terme dans la culture, ce qui permet d’étendre ces tige les populations à volonté pour les applications et les analyses en aval. Pour des raisons inconnues, nous avons échoué à obtenir yMaSC organoïdes en plaçant infecté les cellules directement dans les conditions de culture organoïde LD 7 jours après le traitement doxycycline en plaque de culture de tissu en plastique ; en d’autres termes, l’étape intermédiaire de la croissance dans des conditions de colonie mammaire est indispensable. Dans nos mains, MaSCs même natifs exigent des conditions colonie mammaires avant passage en culture organoïde. En outre, l’excroissance organoïde plus efficace est obtenue quand nous éviter dissociant les colonies primaires dans des cellules isolées, mais plutôt transférer les colonies intactes dans des conditions de culture organoïde.
Conditions de culture organoïde portent également l’avantage de donner la possibilité à cryoconservé CJS, pourvu que les organoïdes sont récupérés à partir de leurs matrices, évitant la dissociation cellulaire avant cryoconservation dans bain d’azote.
Le YAP reprogrammation procédure présentée permet de convertir des types distincts de cellules différenciées provenant de différents tissus adultes dans les cellules correspondantes de souches de tissu-spécifique (nous avons testé à l’aide de cellules mammaires, du pancréas et neuronales)1. À la différence du CISP ou d’autres efforts de reprogrammation, SCs YAP/induite peuvent conserver les souvenirs de leur tissu d’origine. À noter la dédifférenciation des cellules somatiques dans les cellules dotée d’une tige-comme des propriétés est la seule forme de plasticité destin cellulaire et reprogrammation observée in vivo, par exemple après des lésions tissulaires et de soutenir la guérison5,17 , 18 , 19 , 20. il est intéressant de noter que YAP et TAZ sont largement dispensable pour l’homéostasie normale mais fondamental pour la réparation des tissus dans plusieurs tissus11,21. Toujours avec une fonction physiologique des reprogrammation décrites ici, YAP/TAZ a été récemment démontré nécessaires en régénération intestinale chez les modèles murins de patients de la colite ulcéreuse en provoquant une conversion des cellules de l’intestin adultes en un épithélium réparation qui affiche les caractéristiques du tube digestif foetal19. Reprogrammation de YAP élargit ainsi les stratégies actuelles de plasticité cellulaire induite en fournissant un moyen de générer des cellules souches somatiques, un État qui a été jusqu'à présent difficile de capturer in vitro. Cette approche, si étendu aussi aux cellules d’origine humaine, pourrait concernent le vaste d’applications de la médecine régénératrice à l’étude de l’état de stemness somatique et expansion du somatique tige cellules in vitro.
Déclarations de divulgation
Les auteurs déclarent sans intérêts financiers concurrents.
Remerciements
Nous remercions le don de tetO-YAPS127A souris; F. Camargo R26-rtTAM2 souris (stock #006965) ont été achetés chez le laboratoire Jackson. Nous remercions Chiara Frasson et Giuseppe Basso pour obtenir de l’aide des procédures de FACS. Ce travail est soutenu par l’AIRC spécial programme moléculaire Clinical Oncology '' 5 pour mille'' et un PI-Grant AIRC S.P et par le projet phare de l’épigénétique CNR-Miur accorde aux S.P. Ce projet est financé depuis le Conseil européen de recherche (CER) sous recherche Horizon 2020 de l’Union européenne et le programme pour l’innovation (Convention DENOVOSTEM no 670126 de subvention).
matériels
| Name | Company | Catalog Number | Comments |
| 10 mL sterile syringes | Rays | 10LC | |
| 100 mm cell strainer | Corning | 352360 | |
| 15 mL sterile conical tubes | Corning | 430052 | |
| 24-well ultra low attachment plates | Costar | 3473 | |
| 40 mm cell strainers | Corning | 352340 | |
| 48-well multiwell plates | Corning | 353078 | |
| 50 mL sterile conical tubes | Corning | 430290 | |
| 6-well multiwell plates | Corning | 353046 | |
| Advanced DMEM/F12 | Gibco | 12634028 | |
| B27 supplement (50x) | Gibco | 17504001 | |
| BPE | Gibco | 13028014 | |
| BSA | Sigma | A9418 | |
| Collagenase, type I | Sigma | 17018029 | |
| dexamethasone | Sigma | D4902 | |
| Dispase | Gibco | 1705-041 | |
| Disposable scalpels | Swann-Morton | 0503 | |
| DMEM/F12 | Gibco | 11320033 | |
| DMSO | Sigma | D2650 | |
| DnaseI | Roche | 11284932001 | |
| doxycycline hyclate | Sigma | D9891 | |
| EDTA | Sigma | E5134 | |
| Ethanol 100% | Sigma | 51976 | |
| FACS tubes (with strainer caps) | Falcon | 352235 | |
| FBS | Gibco | 10270106 | |
| FITC anti-mouse CD326 (Ep-CAM) | BioLegend | 118208 | |
| FUdeltaGW-rtTA | Addgene | #19780 | |
| FUW-tetO-EGFP | Addgene | #84041 | used as negative control |
| FUW-tetO-MCS | Addgene | #84008 | used as negative control |
| FUW-tetO-wtYAP | Addgene | #84009 | |
| FUW-tetO-YAPS94A | Addgene | #84010 | used as negative control (transcriptionally dead YAP mutant) |
| GlutaMax | Gibco | 35050061 | |
| HBSS | Gibco | 24020117 | |
| HCl | Sigma | 30721 | |
| heparin sodium salt | Sigma | H3149 | |
| HEPES buffer solution (1M) | Gibco | 15630-056 | |
| human R-Spondin1 (His Tag) | Sino Biological | 11083-H08H-5 | |
| Hyaluronidase from bovine testes | Sigma | H3506 | |
| ITS-X | Gibco | 51500056 | |
| K-14 antibody | Life Technologies | Ab7800 | |
| K-8 antibody | Life Technologies | Ab14053 | |
| L-Glutamine | Gibco | 25030081 | |
| Lin (allophycocyanin [APC] mouse lineage antibody cocktail) | BD Biosciences | 51-9003632 | |
| Matrigel® Growth Factor Reduced Basement Membrane Matrix, Phenol Red-Free | Corning | 356231 | |
| N-Acetylcysteine | Sigma | A9165 | |
| NaOH | J.T.Baker | 0402 | |
| NH4Cl | Sigma | A9434 | |
| Nicotinamide | Sigma | 72340 | |
| non-cell adhesive 10 cm dishes (sterile polystirol petri dish ø 94) | ROLL | 18248 | |
| PBS 10x | Euroclone | ECM4004XL | |
| PE Hamster Anti-Mouse CD61 | BD Biosciences | 553347 | |
| PE-Cy5 Rat Anti-Human CD49f | BD Biosciences | 551129 | |
| PE/Cy7 anti-mouse/rat CD29 Antibody | BioLegend | 102222 | |
| Pen/Strep (10,000 U/mL) | Gibco | 15140122 | |
| Rat Tail Collagen I (coating) | Sigma | 122-20 | |
| Rat Tail Collagen I for 3D culture | Cultrex | 3447-020-01 | |
| recombinant human FGF10 | Peprotech | 100-26 | |
| recombinant human Noggin | Peprotech | 120-10C | |
| recombinant murine EGF | Peprotech | 315-09 | |
| recombinant murine FGF basic (bFGF) | Peprotech | 450-33 | |
| RPMI 1640 medium | Gibco | 31870025 | |
| SBTI (Trypsin inhibitor from Glycine max) | Sigma | T6522 | |
| Tris BASE | Roche | 11814273001 | |
| Trypsin-EDTA 0,05% | Gibco | 25300054 | |
| Waymouth medium | Gibco | 31220023 |
Références
- Panciera, T. Induction of Expandable Tissue-Specific Stem/Progenitor Cells through Transient Expression of YAP/TAZ. Cell Stem Cell. 19 (6), 725-737 (2016).
- Bar-Nur, O. Lineage conversion induced by pluripotency factors involves transient passage through an iPSC stage. Nat Biotechnol. 33 (7), 761-768 (2015).
- Xu, J., Du, Y., Deng, H. Direct lineage reprogramming: strategies, mechanisms, and applications. Cell Stem Cell. 16 (2), 119-134 (2015).
- Clevers, H. Modeling Development and Disease with Organoids. Cell. 165 (7), 1586-1597 (2016).
- Blanpain, C., Fuchs, E. Stem cell plasticity. Plasticity of epithelial stem cells in tissue regeneration. Science. 344 (6189), 1242281 (2014).
- Bai, H. Yes-associated protein regulates the hepatic response after bile duct ligation. Hepatology. 56 (3), 1097-1107 (2012).
- Cai, J. The Hippo signaling pathway restricts the oncogenic potential of an intestinal regeneration program. Genes Dev. 24 (21), 2383-2388 (2010).
- Elbediwy, A. Integrin signalling regulates YAP and TAZ to control skin homeostasis. Development. 143 (10), 1674-1687 (2016).
- Taniguchi, K. A gp130-Src-YAP module links inflammation to epithelial regeneration. Nature. 519 (7541), 57-62 (2015).
- Zhang, W. Downstream of mutant KRAS, the transcription regulator YAP is essential for neoplastic progression to pancreatic ductal adenocarcinoma. Sci Signal. 7 (324), ra42 (2014).
- Zanconato, F., Cordenonsi, M., Piccolo, S. YAP/TAZ at the Roots of Cancer. Cancer Cell. 29 (6), 783-803 (2016).
- Azzolin, L. YAP/TAZ incorporation in the beta-catenin destruction complex orchestrates the Wnt response. Cell. 158 (1), 157-170 (2014).
- Stingl, J. Purification and unique properties of mammary epithelial stem cells. Nature. 439 (7079), 993-997 (2006).
- Means, A. L. Pancreatic epithelial plasticity mediated by acinar cell transdifferentiation and generation of nestin-positive intermediates. Development. 132 (16), 3767-3776 (2005).
- Shi, G. Maintenance of acinar cell organization is critical to preventing Kras-induced acinar-ductal metaplasia. Oncogene. 32 (15), 1950-1958 (2013).
- Huch, M. Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 32, 2708-2721 (2013).
- Fernandez Vallone, V. Trop2 marks transient gastric fetal epithelium and adult regenerating cells after epithelial damage. Development. 143 (9), 1452-1463 (2016).
- Yanger, K. Robust cellular reprogramming occurs spontaneously during liver regeneration. Genes Dev. 27 (7), 719-724 (2013).
- Yui, S. YAP/TAZ-Dependent Reprogramming of Colonic Epithelium Links ECM Remodeling to Tissue Regeneration. Cell Stem Cell. , (2017).
- Pan, F. C. Spatiotemporal patterns of multipotentiality in Ptf1a-expressing cells during pancreas organogenesis and injury-induced facultative restoration. Development. 140 (4), 751-764 (2013).
- Panciera, T., Azzolin, L., Cordenonsi, M., Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol. 18 (12), 758-770 (2017).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.