
Method Article
从头用狂吠/狂暴法生成体细胞干细胞
摘要
在再生医学、疾病建模和了解 SC 特性方面, 可获得的躯体 SCs 是至关重要的。在这里, 我们提出的实验策略, 以重新编程,在体外,分化成人细胞成相应的可膨胀的组织特定的茎/祖细胞由单转录联合激活剂狂吠的瞬态表达。
摘要
在这里, 我们提出的协议, 以隔离主要分化细胞, 并把它们变成茎/祖细胞 (SCs) 的同一血统的转录因子狂吠的瞬态表达。采用这种方法, 将小鼠乳腺腔内分化 (LD) 细胞转化为显示乳腺 SCs 分子和功能特性的细胞. 狂吠也将完全分化的胰腺外分泌细胞转变为胰管样祖先。同样, 对内源性的, 自然的 SCs, 狂吠诱导的茎样细胞 ("ySCs") 可以最终扩大为 organoid 文化长期在体外,不需要进一步的异位狂吠/狂暴, 因为 ySCs 被赋予一个可遗传的自我更新 SC 样状态。
这里提出的重新编程程序提供了一种可能性, 以产生和扩大从分化细胞开始的各种组织来源的体外祖细胞。体细胞的直接扩展前体对再生医学有影响, 了解肿瘤起始机制, 更一般的是细胞和发育生物学研究。
引言
组织特异性体细胞干细胞 (SCs) 是损伤后组织更新和修复的关键。容易分离和无限扩展体外体的可能性是潜在再生疗法的一个关键问题, 也是基础研究和疾病建模中的 SC 应用. 然而, 这一方向的进展由于难以捕获各种上皮器官的 SC 状态在体外而受到限制。事实上, 在一些成人组织中, SCs 可能不存在, 或者没有现成的, 或者他们的数量和再生潜能可能因衰老或疾病的情况而受到侵蚀。在 2016年, 我们开始通过报告一个单一转录辅激活因子, 狂吠 (是相关蛋白) 或其密切相关的蛋白 (转录激活物与 PDZ 的基因) 的表达, 以填补这一差距, 到晚期分化细胞有效地创建功能性、可扩展的、非癌变的自体细胞群, 它们与相应的组织特定的 SCs1具有不同的操作性和分子区别。在几天内持续的狂吠或狂暴活动的脉搏足以诱发自我更新的躯体 SCs 的出现。这是一个稳定的条件, 不再依赖于持续的转基因表达, 因为它可以通过细胞世代传播, 而不进一步表达的异位狂吠/狂暴1。本协议详细介绍了从这些组织的分化细胞开始生成乳腺和胰腺的从头上皮茎/祖细胞的程序. 此过程在当前的重新编程/转分化竞技场中填充一个黑色框。这些方向的主要努力实际上已经集中在细胞向诱导多潜能干细胞 (iPSC) 状态的过渡, 然后将这些胚胎和多功能的 SCs 转化为更多的分化细胞。然而, iPSCs 是癌变一旦引入成人组织, 提高了发展协议的需要, 为其完整和有效的分化2。然而, 这种差异化步骤, 即使在可能的情况下, 是以长期可伸缩性、自组织性和器官重写潜能为代价的。这些是器官再生的基本属性, 实际上是典型的唯一的内源性组织特异性 scs 和目前描述的狂吠诱导的 scs (ySCs)。同样, 通过使用各种转录因子的鸡尾酒将一种细胞类型直接转分化为另一种, 也会产生缺乏必要增生和 stemness 潜能的分化细胞3。
这里描述的过程也利用了最近引入的 organoid 技术, 其中内生 SCs 可以扩展和区分前体4。在不存在内源性 scs 的实验、生物学或疾病条件下, 狂吠诱发的 scs 也可能产生 organoid 形成的 scs。我们要指出的是, 在与其他重新编程程序的不同之处, 狂吠所传授的细胞可塑性类型可能对应于活体组织中出现的与 SC 类似的状态的唯一形式。重新取得的特性与组织修复或致癌激活5有关.虽然对几个成人组织的动态平衡, 狂吠和/或狂暴是绝对必要的再生, 肿瘤生长和体外扩张的躯体 SCs1,6,7,8 ,9,10,11,12
研究方案
所有动物程序都遵循我们的制度准则, 并经 OPBA 和卫生部批准。
1. 狂吠诱导的乳腺干样细胞的生成 (yMaSCs)
注意: 1 节的所有媒体和解决方案组合都在表 1中指定。
- 原发性乳腺细胞种群的分离
- 在细胞培养罩下准备: 一次性手术刀, 透明质酸酶溶液, 离解介质, 溶血溶液, 分选溶液, 胶原 i 涂层溶液, Ca2 +螯合液, 洗涤介质 #1, 洗涤介质 #2, 酶溶液,乳房2D 培养基, 冰冷 HBSS/PS。
- 对于一个典型的实验, 牺牲10只雌性小鼠 (CD-1 C57BL/6 菌株), 8-12 周大的颈椎脱位。在解剖前用丰富的70% 乙醇溶液对腹部进行消毒。
- 解剖乳腺通过做一个 Y 形切口沿腹部皮肤和仔细地分离腺体从腹膜通过轻轻地拉扯与。将解剖腺体放置在非细胞粘附盘中, 10 毫升冰冷 HBSS/PS (每道菜20个腺体), 确保不携带任何皮肤碎片。
- 在组织培养罩下, 用10毫升新鲜的 HBSS/PS 洗涤每个腺体, 并将它们放在一个空的非细胞粘附盘中 (每道菜的20个腺体)。不要使用组织培养板, 因为细胞将倾向于坚持他们造成重大损失的材料。
- 用手术刀将乳腺细碎, 直到获得1毫米3片段的均匀混合。用25毫升血清学吸管从每道菜中回收10毫升的分离培养基中的碎组织, 以避免堵塞和转移悬浮在50毫升圆锥管吹打至少 5x, 以分类组织团簇。
注意: 为了有效消化, 在步骤1.1.5 中适当切碎组织是至关重要的。 - 在37摄氏度孵育1小时, 持续剧烈晃动。1小时后, 检查匀浆和延长孵化10分钟, 如果团簇仍然存在。在室温下将所消化的组织在 400 x g 处旋转5分钟, 然后丢弃上清。并用重悬3毫升溶血溶液中的组织颗粒, 在冰上孵化3分钟。
注意: 溶血是一个相当苛刻的治疗, 所以严格的时间是关键的这一步。 - 用10毫升洗涤培养基 #1 冲洗细胞, 将所消化的组织在 400 x g 处旋转5分钟, 然后丢弃上清。并用重悬10毫升的洗涤培养基2和板在10厘米组织培养皿中的组织颗粒。在细胞培养孵化器中孵化1小时37摄氏度的菜肴。
注意: 这一步将允许去除大多数成纤维细胞, 那应该坚持培养皿。 - 从盘子里恢复细胞悬浮液, 倒入50毫升锥形管。旋转 400 x g 5 分钟, 并消除上清。每次在10毫升的 Ca2 +螯合溶液中以 400 x g 为5分钟, 将颗粒清洗两次。并用重悬在5毫升0.25% 胰蛋白酶/EDTA 和孵化5分钟在37摄氏度。
- 在胰蛋白酶溶液的顶端加入5毫升的酶溶液, 并补充 DNase I 1μg/毫升。吸管向上和向下至少5x 通过1毫升尖端, 以分类 DNA 团簇和孵化10分钟37摄氏度, 颤抖每3分钟。
- 添加10毫升的洗涤介质 #2 和过滤器在一个新的50毫升圆锥管通过40微米细胞过滤。将细胞悬浮在 400 x g 处旋转5分钟, 然后丢弃上清, 确保清除所有液体。
- 利用资产管制仪纯化乳腺上皮细胞
- 加入 10 ul 林 (小鼠血统抗体鸡尾酒), 准备抗体组合, 12 ul 反 CD326 (Ep 凸轮), 到最后集中 30 ng/毫升, 10 ul 反 CD49f, 到最后浓度 25 ng/毫升, 10 ul 反 CD61, 最终浓度 10 ng/毫升, 2.5 ul 抗 CD29, 最终浓度为 2.5 ng/毫升。
注意: 从这个步骤到步骤 1.3, 总是在黑暗中操作, 以避免漂白荧光标记的抗体. 对于每个颗粒: - 保持少量的细胞 (通过蘸 100 ul 尖端在小球) 在一个单独的管。并用重悬在 500 ul 的分类溶液中的细胞, 并保持在冰作为一个未标记的样品的外地资产管制程序。
- 并用重悬每个颗粒在 200 ul 的洗涤介质 #1, 添加 44.5 ul 的抗体混合, 吸管彻底和孵化30分钟冰在黑暗中。稀释细胞悬浮在10毫升洗涤介质 #1, 在 400 x g 下旋转5分钟, 并丢弃上清。
- 并用重悬2毫升的分选溶液中的细胞颗粒, 并通过帽过滤器地铁或进行过滤, 进行细胞群分离(如图 1B)。在执行多色的流化程序协议之前, 一定要纠正每个 fluorochrome 的可能溢出。要做到这一点, 每个荧光共轭抗体单独孵化与细胞悬浮和测量光谱重叠值为所有显影和在所有探测器, 通过单色控制, 以创建补偿矩阵。
注: 本实验采用85微米喷嘴的分拣机。来自10只雌性小鼠的典型制剂将产生大约80万个 LD 细胞。进行二次过滤, 如果团簇形式在外地资产管制程序。
- 加入 10 ul 林 (小鼠血统抗体鸡尾酒), 准备抗体组合, 12 ul 反 CD326 (Ep 凸轮), 到最后集中 30 ng/毫升, 10 ul 反 CD49f, 到最后浓度 25 ng/毫升, 10 ul 反 CD61, 最终浓度 10 ng/毫升, 2.5 ul 抗 CD29, 最终浓度为 2.5 ng/毫升。
- 原发性乳腺 LD 细胞的播种
- 在操作过程中, 用胶原 i 涂层溶液涂上多井组织培养板。孵育1小时在37°c, 5% CO2在细胞培养孵化器。取出涂层溶液, 用洗涤介质 #1 电镀前清洗。
- 用10毫升的洗涤液 #1, 将 400 x g 旋转到5分钟, 并消除上清, 从操作过程中恢复的洗涤细胞。并用重悬乳腺2D 培养基中的细胞颗粒 (500 µL/井) 和胶原治疗钢板。让细胞在细胞培养孵化器中坐48小时, 以便适当的细胞附着和传播。
注: 对于10只雌性小鼠的典型排序, 24 井多井板6-8 井将产生最佳细胞密度 (100 000 细胞/井)。
- yMaSCs (狂吠诱导的乳腺干细胞) 的诱导
注意: 从这一步开始, 所有程序都应在 BSL-2 条件下执行。- 准备乳房菌落培养基。刚在细胞播种前将基底膜基质添加到培养基中。
- 对狂吠诱导的乳腺干细胞 (yMaSCs) 的诱导, 传感器慢病毒载体感染的主要 LD 细胞, 混合一量 FUdeltaGW rtTA 病毒上清, 一容积的福网-tetO-狂吠 (或狂暴) 上清, 两卷无血清乳房2D 培养基2x 浓度的补充剂, 总体积为500毫升。用慢病毒载体上清液孵化细胞48小时。对于典型的慢病毒载体准备, 请参阅联机协议22。
- 感染后, 用2µg/毫升强力霉素, 用乳腺2D 培养基对黏附细胞进行冲洗, 以诱导外源性狂吠 (或狂暴) 基因表达。使用细胞感染的空, EGFP 或 YAPS94A 表达载体, 或细胞感染诱导狂吠 (或狂暴) 的载体, 但左无强力霉素作为阴性对照。
注意: 成功的感染可以通过 qRT PCR 方法来验证, 如前所述的1, 它是针对人狂吠转基因的底漆。 - 经过7天的诱导与强力霉素, 分离黏附细胞通过孵化与0.05% 胰蛋白酶/EDTA (150 ul/井) 10 分钟37°c;停止 trypsinization 稀释1:5 在洗涤介质 #2 (600 ul/井) 和计数细胞。并用重悬细胞在乳腺菌落培养基 (每井1毫升), 补充2微克/毫升强力霉素和种子在克隆密度1000细胞/井在24以及超低的附件板。
注意: 在基底膜基质添加时, 要确保乳腺菌落培养基为冰寒。基底膜基质在到达时必须始终保存在-20 摄氏度, 并在一夜之间慢慢解冻4摄氏度;一旦解冻, 它必须总是在冰上处理, 根据制造商的指导方针。 - 一旦狂吠表达 LD 细胞开始增殖和成长为 MaSC 样的殖民地在悬浮 (yMaSC 殖民地) (播种后14天), 计数和进程进一步分析 (图 1C)。
注意: 负控制单元 (如点 1.4.3) 将保持为单个细胞。 - 在 yMaSC 菌落生长14天内, 每72小时补充新鲜乳房菌落培养基的培养;要做到这一点, 准备一个整除的乳房菌落培养基没有5% 基底膜基质, 补充10x 浓度的补充剂, 并增加1:10 的总体积每井 (例如100 ul 在1毫升的总培养基), 以避免过度稀释矩阵悬浮。
- yMaSCs 的亚培养
- 从乳腺菌落培养基中恢复主要菌落, 然后分离和补种。
注: 由狂吠重组的 LD 细胞衍生的 yMaSC 菌落获得自我更新能力, 如果没有进一步的强力霉素管理 (i. e, 独立于转基因吠/狂暴的表达), 可以成功地进行亚培养。 - 准备, 在细胞培养罩下, 乳腺菌落培养基为阶梯1.4.1 和乳腺 organoid 培养基。
- 收集每个样品和孵化在多余的体积 (10:1) 冰冷 HBSS 1 小时冰, 以溶解基底膜基质。用离心在 180 x g 5 分钟和并用重悬冰 HBSS 中清洗菌落3x。孵化菌落在0.05% 胰蛋白酶/EDTA 10 分钟37°c, 以获得一个单一的细胞悬浮。用 p1000 尖吸出10x 的吸管, 确保完全离解到单细胞水平。
- 计数和补种细胞在乳房殖民地媒介 (每井1毫升), 不用强力霉素在克隆密度1000个细胞/井在24和超低的附件板材。每10-14 天重复这个传代程序来评估自我更新。
- 在第三段之前, 通过 yMaSC 殖民地在 organoid 文化条件, 加强 yMaSC 扩展和允许形成微型腺体, 那自我组织在双层上皮密切地令人想起在体内乳腺组织学组织。
- 从乳腺菌落培养基中恢复菌落, 如步1.5.3 到1.5.4。并用重悬在100% 生长因子中的菌落减少了基底膜基质, 考虑到在基体的150µL 中, replate 24 井超低附着板的每个井的最大20-25 个菌落。
- 在37摄氏度的细胞培养孵化器中孵化出40分钟的板块, 然后让基底膜基质凝固, 然后用乳腺 organoid 培养基的 500 ul 对凝胶进行叠加。
- 几天后, 检查形成的殖民地, 形成萌芽 organoids (图 1E)。
- 经过10-14 天, 通过或处理 organoids 进行进一步分析。
- 为了通过 organoids 文化, 通过收集每个样品和孵化在10:1 冰冷 HBSS 1 小时在冰, 为了溶解基底膜基质, 恢复 organoids。用 180 x g organoids 5 分钟, 并用重悬冰 HBSS, 洗净3x。
- 孵育 organoids 在0.05% 胰蛋白酶/EDTA 10 分钟37°c, 以获得一个单一的细胞悬浮。吸管 organoids 向上和向下10x 与 p1000 提示, 以确保完全离解到单细胞水平。
- 补种作为单细胞悬浮在一滴100% 生长因子减少基底膜基质 (150 ul 为每井24井超低附着板)。让基底膜基质形成一个凝胶通过孵化40分钟在37°c 在细胞培养孵化器和覆盖凝胶与 500 ul 的乳房 organoid 培养基。
注: yMaSC organoids 可通过从100% 基底膜基质培养中恢复为步进 1.5.13, 避免 trypsinization。贮存在乳腺 organoid 培养基中, 辅以10% 亚砜。 - 快速冷冻 yMaSC organoids-80 摄氏度, 然后保存在液氮中。
- 从乳腺菌落培养基中恢复主要菌落, 然后分离和补种。
2. yDucts 的产生
注意: 2 节的所有媒体和解决方案组合都在表 2中指定。
- 原发性胰腺腺泡细胞的分离
- 放置解剖钳和剪刀在70% 乙醇和准备下细胞培养罩腺泡培养培养基, 15 毫升为每只老鼠;腺泡恢复培养基, 每只老鼠60毫升;PBS/PS;库存溶液胶原酶 I;胶原酶 I 溶液 A, 每只老鼠15毫升;中性鼠尾胶原蛋白 i 溶液。中和鼠尾胶原蛋白 i 到 pH 值 = 7, 先用0.1 氮氢氧化钠来调节胶原溶解的醋酸, 再用 10N HCl. 在 PBS/PS 中稀释至2.5 毫克/毫升. 保持鼠尾胶原蛋白 i 和所有的试剂在冰上中和。牺牲6到9周--正常基因型的老鼠。
- 将每只老鼠放在背上, 用70% 乙醇溶液冲洗腹部。沿腹壁进行纵切口。定位和解剖胰腺 (以脾脏为导向), 并将其放置在10厘米非细胞粘附盘中, 10 毫升冰冷 PBS/PS. 立即将盘子转移到细胞培养罩下。从这个步骤开始, 工作总是在细胞培养罩下。
- 转移每个胰腺在一个新的非细胞粘附盘以前填充7毫升胶原酶 I 溶液 a。
- 用一双一次性手术刀快速切碎每个胰腺, 以获得大约1毫米3片段的均匀组织悬浮。
注意: 重要的是要注意, 这一过程应采取不超过2分钟的最佳细胞活力。 - 为胶原酶消化37摄氏度, 5% CO2在细胞培养孵化器中孵育10分钟, 每3分钟晃动, 以保证均匀的组织消化。
- 恢复50毫升圆锥管 (每一个胰腺) 的消化组织, 用10毫升的腺泡洗涤培养基冲洗盘子, 将其放在同50毫升圆锥管中, 吹打消化组织不超过3x。
- 在18摄氏度的 100 x g 处旋转消化组织5分钟, 取出上清液。
注: 在这个步骤中, 在18摄氏度以下的细胞向下旋转以降低胶原酶活性。 - 并用重悬7毫升胶原酶 I 溶液中的组织颗粒, 并将此溶液倒入新的10厘米非细胞胶盘中。
- 孵育第二轮胶原酶消化37°c 为10分钟, 在步骤 2.1.5, 摇晃每3分钟, 以确保均匀组织消化。同时, 为每个胰腺准备一个干净的50毫升圆锥管, 上面有100微米细胞过滤器。
- 恢复消化组织和通过100微米细胞过滤器通过浸泡的组织与无菌10毫升注射器柱塞 (确保仔细压下组织, 避免剪切力切线到过滤器表面)。用10毫升的腺泡洗涤培养基冲洗盘子, 通过同一100微米细胞过滤器将这10毫升通过。
- 在18摄氏度的 100 x g 处旋转消化组织5分钟, 取出上清液。
- 用10毫升腺泡洗涤培养基回收组织颗粒。在50毫升圆锥管中转移细胞溶液, 其中含有额外的10毫升新鲜的腺泡洗涤培养基, 避免过量吹打泥沙的颗粒。
- 在18摄氏度的 100 x g 处旋转消化组织5分钟, 取出上清液。
- 仔细并用重悬6毫升腺泡回收培养基中的消化组织, 并将其分布在2口6井多井组织培养板, 每3毫升。在显微镜下, 仔细评估腺泡分离的质量, 它表现为腺泡簇的均匀悬浮, 单个细胞比例较小 (参见图 2B);去除任何大的组织团簇最终呈现 (通常也可见肉眼), 通过吹打他们的解决方案。
- 原发性胰腺腺泡细胞的播种
- 在细胞培养孵化器中孵育37摄氏度的消化腺泡簇, 以使细胞恢复。
- 在细胞恢复大衣48井多井与 100 ul 的中和大鼠尾胶原蛋白 I 和孵化1小时37°c 在细胞培养孵化器, 以允许水凝胶垫形成。
- 经过2小时的细胞恢复, 收集腺泡细胞悬浮在锥形管, 旋转下来5分钟 100 x g 在18°c, 并删除上清。
- 并用重悬腺泡细胞在腺泡培养基适当容积 (150 ul 为48井组织培养板材的每个井)。将每个分离的胰腺种子分成16口, 获得最佳密度 (100-120 腺泡簇/井)。
- 用等量的中和大鼠尾胶原 i 溶液稀释这个腺泡悬浮液, 把管子放在冰上。仔细和快速地混合在2.2.2 描述的胶原垫上的细胞悬浮液 (48 井多井组织培养板每井 300 ul)。
- 孵育1小时在37°c 在细胞培养孵化器允许水凝胶形成。
- 胰腺 organoids 的诱导
- 用 500 ul 腺泡培养培养基覆盖胶原凝胶, 辅以2微克/毫升强力霉素, 用于从 R26-rtTAM2 诱导胰 organoids ;TetO-狂吠S127A 鼠标;负控制是由在相同条件下培养的 R26-rtTAM2 单元提供的, 或者是 。TetO-狂吠S127A 在没有强力霉素的情况下培养的细胞。
- 腺泡培养培养基中培养的腺泡细胞, 辅以2微克/毫升强力霉素5至7天清爽培养基 (300 ul/井) 每 48 h 和以下 organoid 形成由形态学变化对囊肿形成导管样结构 (图 2C)。一旦 organoids 形成, 细胞可以传代胰腺 organoid 培养条件或收获进一步分析 (例如: RNA 提取, 免疫荧光)。
- 胰 organoids 的亚培养
- 为了评估其自我更新能力, 无性通过狂吠诱导胰腺 organoids (yDucts) 在三维基底膜基质水凝胶 (胰腺 organoid 培养条件) 独立的外源吠/狂暴供应 (i. e。, 独立于强力霉素管理)。
- 制备胰蛋白酶 005%/EDTA;100% 生长因子降低基底膜基质;胰 Organoid 培养基和胶原酶 I. 的溶液 B。
- 准备一个15毫升圆锥管与4毫升胶原酶 I 溶液 B 为每个井被传代。
- 摒弃培养基, 小心翼翼地从水井中提取水凝胶, 并将其转移到锥形管上。
- 孵化管在37°c 30 分钟, 持续剧烈晃动, 允许完全消化胶原基质 (检查每10分钟, 直到水凝胶完全可溶性)。在 750 x g 处向下旋转恢复的细胞2分钟, 并清除上清。
- 在1毫升的胰蛋白酶 005%/EDTA 中孵育恢复细胞, 在37摄氏度10分钟, 以获得单个细胞悬浮。稀释胰蛋白酶与9毫升 PBS 1x, 旋转下来在 750 x g 为2分钟和去除上清。
- 并用重悬在冰冷生长因子中的细胞颗粒减少基底膜基质和种子在超低附着板 (通常是一滴 150 ul 在一个井的24井板)。
- 让基底膜基质水凝胶通过孵化细胞培养孵化器中的板块40分钟在37摄氏度, 然后覆盖与胰腺 Organoid 培养基 (每井 500 ul) 固化。yDucts 将在7-10 天内成长为囊肿样 organoids (图 2D)。
- 为进一步传代 organoids 可以从地下室膜基质中通过孵化在冰冷的 pbs 1x 30 分钟, 然后洗3次, 在 180 x g 向下旋转5分钟和泥沙在冰冷 pbs 1x, 以避免矩阵的结转。然后将 Organoids 与胰蛋白酶0.05% 分离, 以10分钟的 reseeded 在新鲜基底膜基质中获得单细胞悬浮液, 然后用胰 Organoid 培养基 (如 2.4. 7-2. 4.8) 进行叠加。
- yDuct organoids 可在液氮中低温保存, 从100% 基底膜基质培养中恢复为步向 2.4.9, 避免 trypsinization, 并以10% 亚砜为胰 Organoid 培养基。
- yDuct organoids 在-80 ºC 迅速冻结, 然后保存在液氮中。
结果
yMaSCs 的产生
在图 1A中介绍了用狂吠瞬时表达重新编程原发性乳腺 LD 细胞的实验策略。原发性乳腺 LD 上皮细胞由荧光活化细胞分类13纯化。图 1B表示一个典型的排序过程, 以获得三个不同的亚群: 基底细胞 (EpCAM低CD49f高CD61), 腔内祖 (LP) 细胞 (EpCAM高CD49f低CD61+) 和 LD 单元格 (EpCAM高CD49f低CD61).仔细浇三亚人口是必不可少的, 以隔离的纯制备 LD 细胞, 这是完全区分和完全增长被捕时, 种子在乳腺菌落形成条件 (见图 1C, 左面板)。相反, 当诱导表达外源狂吠时, LD 细胞开始增殖, 形成容易辨认的密集上皮菌落在5% 基底膜基质悬浮培养 (图 1C)。重新编程的效率, 证实了大约3% 为一个典型的实验, 可以通过计数殖民地数量在最初播种在基底膜基质悬浮培养的单细胞数量 (图 1D)。重新编程的腔细胞 (yMaSCs) 可以通过进入100% 基底膜基质 organoid 培养条件 (参见图 1A中的方案), 自组织成复杂的 organoid 状结构, 围绕多个流明发展, 并即使没有强力霉素 (即缺乏转基因狂吠表达), 也显示出卓越的自我更新能力 (图 1E)。组织学上, yMaSC 衍生 organoids 显示基底层 (K14 阳性), 面对基底膜基质重组 ECM 和一个腔层 (K8 阳性), 面对在 organoid 内的腔样腔 (图 1F)。此体系结构与本机 MaSCs 形成的 organoids (图 1F) 不区分。
yDucts 的产生
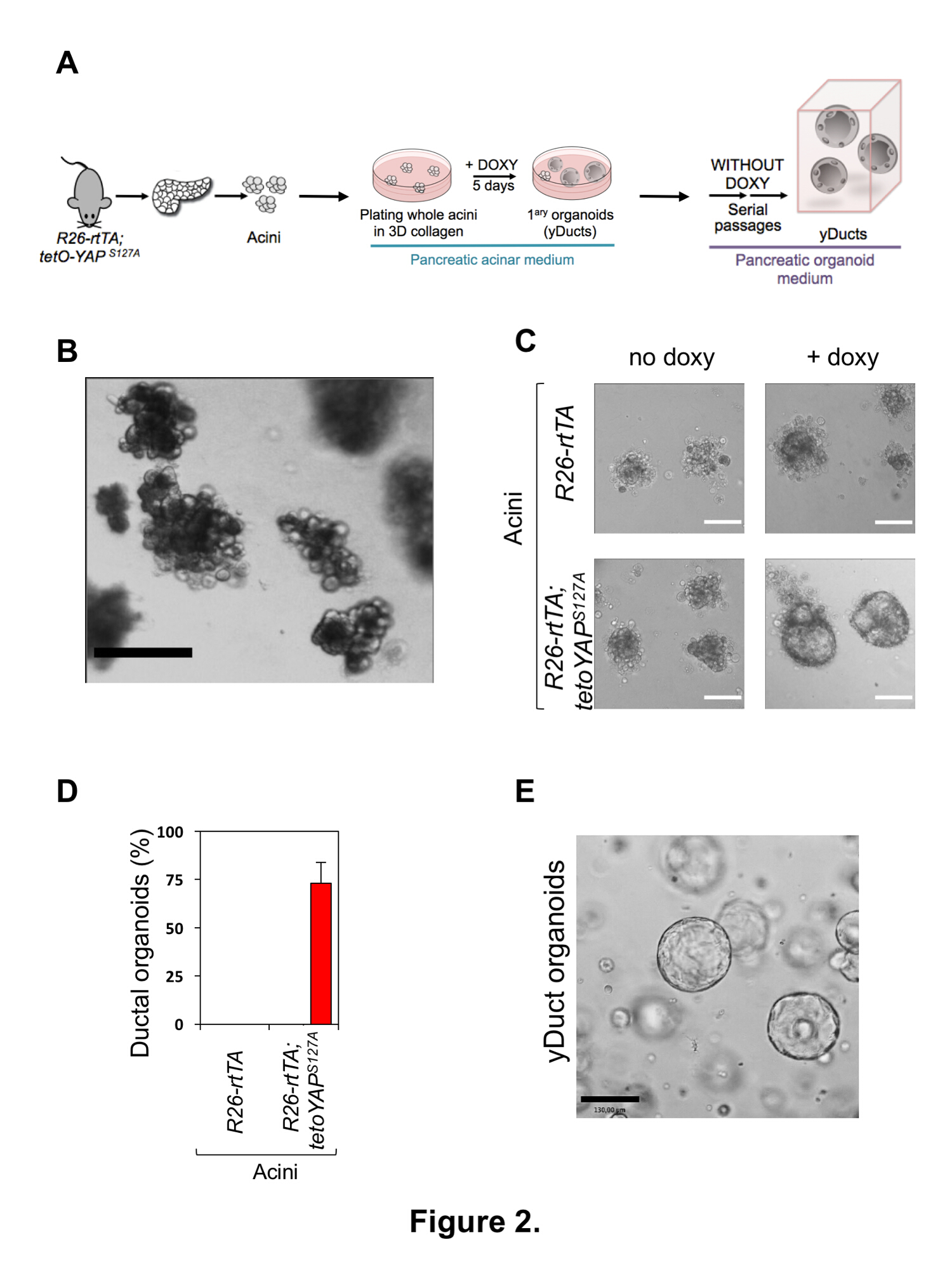
在图 2A中, 概述了用狂吠的瞬态表达重新编程原发性胰腺腺泡细胞的实验策略。整个腺泡簇与胰腺组织的大部分分离, 通过过滤的轻度离解和大小排斥相结合。典型的准备工作在图 2B中显示。分离后, 腺泡细胞簇应表现为一个悬浮的外分泌腺泡单位的均匀大小, 没有污染的内分泌胰岛或片段的胰导管树和最小离解的单一细胞。内分泌胰岛或导管碎片的污染是指缺乏选择性过滤 (步骤 2.1.10), 可能是由于苛刻的处理;腺泡簇与单个细胞的不想要的分离可能是由于过度的胶原酶治疗或组织释放的蛋白水解酶的缓冲活性, 可以通过额外的 SBTI 治疗来抑制。
在图 2C中提出了一种典型的腺泡重编程实验;在5-7 天的培养3D 胶原蛋白-I 基水凝胶在存在强力霉素, 胰腺腺泡细胞从R26-rtTAM2/tetO-YAP S127A 小鼠容易地变成管道状簇 (我们命名yDucts), 由一个薄上皮细胞在扩张中心腔周围增殖的单层。对于一个典型的实验来说, 重新编程效率大约是 70%, 可以通过在种子腺泡细胞的总数上评分像管状簇的数量(图 2D) 来轻松地测量。负控制单元, 即R26-rtTAM2/+单元格或R26-rtTAM2/tetO-YAPS127A 单元格没有强力霉素, 在这些文化条件下始终保持为有丝分裂后腺泡簇, 如先前报告的1 ,14,15。然后, 重新编程的 yDucts 可以在单个单元格级别传代为基于胶的 organoid 区域性条件16 (参见图 2A中的方案), 即使在没有强力霉素的情况下也显示出显著的自我更新能力 (即没有转基因狂吠表达) (图 2E)。

图 1:原发性乳腺 LD 细胞的分离和乳腺干细胞的诱导。(A)用于重新编程初级乳腺 LD 细胞的实验程序的示意图表示法。(B) 有代表性的外地资产管制署--说明典型的分拣程序以净化 LD 细胞的地块。i) 分离细胞根据正向和侧向散射对活细胞进行门控 (P1; 蓝色);二) 然后根据其林廓线进一步封闭人口 P1: 血统阴性细胞亚群 (P2; 灰色) 被选择, 不包括谱系阳性造血细胞;iii) 人口 P2 然后被分离成一个 EpCAM高(P3; 黄色 + 绿色) 和 EpCAM低(P6; 红色) 亚群;iv) P3 和 P6 然后进一步被门控根据他们的 CD61/CD49f 外形成三个亚人口: EpCAM低CD49f高CD61-基底细胞 (P7; 红色), EpCAM高CD49f低CD61+ LP 单元格 (P8; 黄色)和 EpCAM高CD49f低CD61 LD 单元格 (P9; 绿色). (C)图像说明了在乳腺菌落培养基中播种15天后, LD 细胞感染了所述结构, 形成乳腺菌落的能力。只有狂吠表达的细胞变成成藏细胞, 而负控制细胞 (EGFP 感染) 保持为生长被捕获的单个细胞。刻度条 = 50 微米。(D)对所示细胞的菌落形成能力进行量化, 如 (C)。数据显示为平均 + s.d., 并代表五独立实验, 每一个都有六技术复制。(E)在不存在强力霉素的情况下, 在12天后, 在新鲜三维100% 基底膜基质水凝胶中 organoid 培养条件下, 以狂吠重组的乳腺干细胞样细胞 (yMaSCs) 为代表的图像。刻度条 = 100 微米。(F)具有代表性的免疫荧光图像, 用于基底标记 K14 (绿色) 和 organoids 的腔内标记 K8 (红色), 经过12天的 organoid 培养条件。刻度条 = 10 微米。此数字从 Panciera et . 20161中复制。请单击此处查看此图的较大版本.
{kind=link}

图 2: 原发胰腺腺泡细胞的分离及胰祖的诱导。(A)对原发性胰腺外分泌腺泡细胞重新编程所采用的实验程序的示意图表示。(B)仅在隔离过程之后 (步骤 2.1.14) 的原发性胰腺腺泡细胞的代表性图像。腺泡的制备应表现为腺泡簇的均匀悬浮, 单细胞很少存在。刻度条 = 400 微米。(C)从R26-rtTAM2 (上部面板) 或 R26-rtTAM2 中提取的原发性胰腺腺泡细胞的代表性图像; tetO-狂吠 S127A (下板) 小鼠和培养在3维胶原 i 基水凝胶5天有或不强力霉素 (教义), 如所示。在强力霉素添加后, 只有狂吠表达的原发性腺泡细胞转化为像囊肿样 organoid 生长的细胞。刻度条 = 70 微米。(D)对胰腺腺泡细胞形成导管 organoids 的能力进行量化, 如 (C) 所表达的转基因狂吠。数据显示为平均 + s.d., 并代表五独立实验, 执行四技术复制。(E) reprogramed 导管样细胞 (yDucts) 在三通道后在新鲜三维100% 基底膜基质水凝胶中的代表性图像, 不存在强力霉素。刻度条 = 130 微米。请单击此处查看此图的较大版本.
{kind=link}
| 原发性乳腺细胞的分离 | |
| Ca2 +螯合解决方案 | 存储在4°c |
| Edta | 0.02% 瓦特/V |
| Pbs | |
| 胶原 i 涂层溶液 | |
| 乙酸 0.02N, pH 值323 | |
| 鼠尾胶原蛋白 (涂层) | 1:50 |
| 酶解决方案 | 存储在4°c |
| 酶 | 5毫克/毫升 |
| Pbs | |
| 离解介质 | |
| DMEM:F12 | |
| 透明质酸酶库存解决方案 | 400 U/毫升 |
| 笔/链球菌 | 1x |
| 库存溶液胶原酶 I | 600 U/毫升 |
| 溶血解决方案 | 存储在4°c |
| NH4Cl 解决方案 | 1部分 |
| TrisBase 20.6 克/升 | 9部分 |
| 将 pH 值调整为7。2 | |
| HBSS/PS | 存储在4°c |
| HBSS | |
| 笔/链球菌 | 2x |
| 透明质酸酶库存解决方案 | 过滤器0.2 µm, 存储在4°c |
| 牛睾丸透明质酸酶 (粉末) | 2000 U/毫升 |
| 磷酸钠缓冲液 1M pH7。3 | |
| NH4Cl 解决方案 | 存储在磁悬浮轴承上。 |
| H2O | |
| NH4Cl | 7.1 克/升 |
| 将 pH 值调整为7.65 | |
| 排序解决方案 | 过滤器0.2 µm, 存储在4°c |
| Bsa | 0.1% |
| Edta | 1毫米 |
| HEPES pH 值7 | 25毫米 |
| Pbs | |
| 洗涤介质 #1 | |
| DMEM/F12 | |
| 笔/链球菌 | 1x |
| 洗涤介质 #2 | |
| DMEM/F12 | |
| FBS | 5% |
| 笔/链球菌 | 1x |
| 乳腺2D 培养基 | |
| DMEM/F12 | |
| FBS | 2% |
| 肝 素 | 4毫克/毫升 |
| l-谷氨酰胺 | 1x |
| 小鼠 bFGF | 10 ng/毫升 |
| 小鼠 EGF | 10 ng/毫升 |
| 笔/链球菌 | 1x |
| yMaSCs 的诱导与传代 | |
| 乳腺菌落培养基 | |
| DMEM:F12 | |
| FBS | 5% |
| 肝 素 | 4µg/毫升 |
| l-谷氨酰胺 | 1x |
| 胶 (播种前立即添加) | 5% |
| 小鼠 bFGF | 20 ng/毫升 |
| 小鼠 EGF | 10 ng/毫升 |
| 笔/链球菌 | 1x |
| 乳腺 Organoid 培养基 | |
| 高级 DMEM:F12 | |
| B27 | 1x |
| GlutaMax | 1x |
| 肝 素 | 4µg/毫升 |
| Hepes | 1x |
| 人的脑袋 | 100 ng/毫升 |
| 小鼠 bEGF | 20 ng/毫升 |
| 小鼠 EGF | 50 ng/毫升 |
| R-Spondin 1 | 1µg/毫升 |
表 1: yMaSCs 的生成.所有不同培养基的组成和分离原发乳腺 LD 细胞和诱导 yMaSCs 的解决方案 (1 节)。
| 原发性胰腺腺泡细胞的分离 | |
| 腺泡培养基 | |
| Bpe | 50µg/毫升 |
| Bsa | 0.1% |
| 地 塞 米松 | 1µg/毫升 |
| FBS | 0.1% |
| 其 x | 1x |
| 笔/链球菌 | 1x |
| SBTI | 0.2 毫克/毫升 |
| Waymouth 的中等 | |
| 腺泡洗涤培养基 | |
| Bsa | 0.1% |
| 笔/链球菌 | 1x |
| RPMI 介质 | |
| SBTI | 0.2 毫克/毫升 |
| 腺泡回收培养基 | |
| 腺泡培养基 | |
| FBS | 30% |
| 胶原酶 I 溶液 A | |
| 腺泡洗涤培养基 | |
| 库存溶液胶原酶 I | 360 U/毫升 |
| PBS/PS | 存储在4°c |
| PBS (磷酸盐缓冲盐水) | |
| 笔/链球菌 | 1x |
| 库存溶液胶原酶 I | 存储在-20 °c |
| I 型胶原酶 (粉末) | 6000 U/毫升 |
| Pbs | |
| 胰腺 organoids 的传代 | |
| 胶原酶 I 溶液 B | |
| PBS 1x | |
| 库存溶液胶原酶 I | 240 U/毫升 |
| 胰 Organoid 培养基 | |
| 高级 DMEM/F12 | |
| B27 | 1x |
| 胃 泌 素 | 10毫微米 |
| 人类 FGF10 | 100 ng/毫升 |
| 人的脑袋 | 100 ng/毫升 |
| 小鼠 EGF | 50 ng/毫升 |
| n-乙酰半胱氨酸 | 1.25 毫米 |
| 烟 酰 胺 | 10毫米 |
| 笔/链球菌 | 1x |
| R-Spondin 1 | 1毫克/毫升 |
| SBTI | 0.2 毫克/毫升 |
表 2: yDucts 的生成.所有不同培养基的组成以及分离原发泡细胞所需的溶液和 yDucts 的诱导和传代 (2 节)。
讨论
在这里, 我们提出的协议, 重新编程的前体晚期分化上皮细胞的不同组织到其相应的组织特异性祖细胞 (或 ySCs) 的瞬态表达的狂吠, 如报告前1。我们有详细的两个程序: 一个允许重新编程的慢病毒载体载体纯化细胞和第二个避免病毒感染, 并利用转基因狂吠表达。每个协议都提出了一个有效的策略, 分离和培养的主要分化细胞和策略, 以迫使外源狂吠基因表达的分化细胞, 产生从头体细胞组织特定的可膨胀干细胞 (见图 1A和2A中的方案。
我们证明, 这里的隔离策略有效地隔离了一个纯的分化细胞群体, 这是因为我们从来没有从阴性控制样本 (图 1C和2C) 中检测出任何结果。
本研究中使用的慢病毒载体载体重编程的原发性乳腺 LD 细胞是强力霉素诱导, 提供了严格控制转基因表达的可能性;这允许打开和关闭外源狂吠表达的意愿。应特别注意避免使用过量的病毒滴度, 因为这在重新编程效率方面可能有害。在原发性腺泡细胞的情况下, 我们改用完全转基因的方法来获得一项与狂吠相关的重新编程和最小的操作。后者的策略也特别适合于原发性胰腺腺泡细胞, 因为孤立的腺泡群几乎不能适应慢病毒载体感染, 非常脆弱。所采用的转基因策略提供了与强力霉素依赖的慢病毒载体载体相同的优势, 以严格控制基因表达。此外, 与病毒诱导的乳腺 LD 细胞再编程相比, 利用原代胰腺腺泡细胞的转基因策略具有更高的重编程效率。除了与来自不同组织的细胞不同的内在可塑性外, 胰腺重编程的速度可能来自于所有说明细胞。值得注意的是, 我们已经证明了在 ySCs (yMaSC 殖民地和 yDucts) 之后, 外源狂吠不再需要, 而不影响他们的自我更新能力。这是因为 ySCs 重新激活内生吠/狂暴, 并使用它们进行自我更新时, 外源狂吠关闭1。
我们验证了这种观点, 即 ySCs 确实是通过遗传沿袭-跟踪验证1控制我们重新编程实验的起源单元来从分化的细胞中出现的。
ySCs 的广泛特性表明, 狂吠诱导的重编程生成正常的躯体 SCs1作为 i) 在 transcriptomic 水平, ySCs 显示大量重叠与当地的 SCs;ii.) ySCs 显示分化潜能, 并能产生多向的后代, 始终限于其起源组织的身份;iii) 在移植体内时, ySCs 是非变换的和非癌变的。
在这里, 我们还描述了保持和扩大在文化中 yMaSCs 和 yDucts 作为 organoids 嵌入100% 基底膜基质水凝胶的过程。这些条件允许将 ySCs 自组织成三维 organoids, 确保在文化中长期保持 stemness 性质, 从而能够将这些茎群扩大到下游分析和应用。由于不确定的原因, 我们未能获得 yMaSC organoids 通过放置感染的 LD 细胞直接在 organoid 培养条件下, 强力霉素治疗后7天的塑料组织培养板;换言之, 乳腺菌落条件中的中间生长步骤是必不可少的。在我们的手中, 即使是本土的 MaSCs 需要在 organoid 文化传代之前的乳房殖民地条件。此外, 最有效的 organoid 产生是当我们避免游离的主要殖民地的单细胞, 而是把完整的殖民地转移到 organoid 的文化条件。
Organoid 的培养条件也具有 cryopreserve ySCs 的优势, 前提是 organoids 从母体中恢复, 避免在氮气浴保存前细胞离解。
所提出的狂吠重新编程程序可以将不同的成人组织产生的不同的分化细胞类型转化为相应的组织特异干细胞 (我们用乳腺、胰腺和神经细胞进行了测试)1。与 iPSCs 或其他重新编程的努力不同, 狂吠/诱导的 SCs 可以保持他们的起源组织的记忆。注意, 体细胞分化为具有茎样性质的细胞, 是细胞命运可塑性和重新编程在体内观察的唯一形式, 例如组织损伤和支持伤口愈合5,17,18,19,20. 值得注意的是, 狂吠和狂暴基本上是可有可无的正常动态平衡, 但关键的组织修复在多个组织11,21。与这里描述的重新编程步骤的生理功能一致, 最近表明, 在溃疡性结肠炎患者的小鼠模型肠道再生需要的吠/狂暴, 导致成人肠道细胞转化为用于显示胎儿肠道特征的修复上皮19。吠说重新编程从而扩大了目前诱导细胞可塑性的策略, 提供了一种方法来产生体细胞, 这个国家已经到目前为止具有挑战性的捕获在体外。这种方法, 如果也扩展到人类衍生细胞, 可能具有广泛的相关性, 从再生医学应用到研究的躯体 stemness 状态和扩展的体细胞干细胞在体外。
披露声明
作者声明没有竞争的财政利益。
致谢
感谢 f. Camargo 为tetOS127A 小鼠提供的礼物;R26-rtTAM2老鼠 (股票 #006965) 是从杰克逊实验室购买的。我们感谢 Chiara Frasson 和他的低音帮助与外地资产管制程序。这项工作由 AIRC 特别计划分子临床肿瘤学 "5 每比" 和 AIRC PI 补助金对 S P 和由表观遗传学旗舰项目 CNR-Miur 赠款某地该项目已获得欧洲研究理事会 (紧急救济委员会) 根据欧洲联盟2020研究和创新方案 (赠款协议 DENOVOSTEM 670126) 提供的资金。
材料
| Name | Company | Catalog Number | Comments |
| 10 mL sterile syringes | Rays | 10LC | |
| 100 mm cell strainer | Corning | 352360 | |
| 15 mL sterile conical tubes | Corning | 430052 | |
| 24-well ultra low attachment plates | Costar | 3473 | |
| 40 mm cell strainers | Corning | 352340 | |
| 48-well multiwell plates | Corning | 353078 | |
| 50 mL sterile conical tubes | Corning | 430290 | |
| 6-well multiwell plates | Corning | 353046 | |
| Advanced DMEM/F12 | Gibco | 12634028 | |
| B27 supplement (50x) | Gibco | 17504001 | |
| BPE | Gibco | 13028014 | |
| BSA | Sigma | A9418 | |
| Collagenase, type I | Sigma | 17018029 | |
| dexamethasone | Sigma | D4902 | |
| Dispase | Gibco | 1705-041 | |
| Disposable scalpels | Swann-Morton | 0503 | |
| DMEM/F12 | Gibco | 11320033 | |
| DMSO | Sigma | D2650 | |
| DnaseI | Roche | 11284932001 | |
| doxycycline hyclate | Sigma | D9891 | |
| EDTA | Sigma | E5134 | |
| Ethanol 100% | Sigma | 51976 | |
| FACS tubes (with strainer caps) | Falcon | 352235 | |
| FBS | Gibco | 10270106 | |
| FITC anti-mouse CD326 (Ep-CAM) | BioLegend | 118208 | |
| FUdeltaGW-rtTA | Addgene | #19780 | |
| FUW-tetO-EGFP | Addgene | #84041 | used as negative control |
| FUW-tetO-MCS | Addgene | #84008 | used as negative control |
| FUW-tetO-wtYAP | Addgene | #84009 | |
| FUW-tetO-YAPS94A | Addgene | #84010 | used as negative control (transcriptionally dead YAP mutant) |
| GlutaMax | Gibco | 35050061 | |
| HBSS | Gibco | 24020117 | |
| HCl | Sigma | 30721 | |
| heparin sodium salt | Sigma | H3149 | |
| HEPES buffer solution (1M) | Gibco | 15630-056 | |
| human R-Spondin1 (His Tag) | Sino Biological | 11083-H08H-5 | |
| Hyaluronidase from bovine testes | Sigma | H3506 | |
| ITS-X | Gibco | 51500056 | |
| K-14 antibody | Life Technologies | Ab7800 | |
| K-8 antibody | Life Technologies | Ab14053 | |
| L-Glutamine | Gibco | 25030081 | |
| Lin (allophycocyanin [APC] mouse lineage antibody cocktail) | BD Biosciences | 51-9003632 | |
| Matrigel® Growth Factor Reduced Basement Membrane Matrix, Phenol Red-Free | Corning | 356231 | |
| N-Acetylcysteine | Sigma | A9165 | |
| NaOH | J.T.Baker | 0402 | |
| NH4Cl | Sigma | A9434 | |
| Nicotinamide | Sigma | 72340 | |
| non-cell adhesive 10 cm dishes (sterile polystirol petri dish ø 94) | ROLL | 18248 | |
| PBS 10x | Euroclone | ECM4004XL | |
| PE Hamster Anti-Mouse CD61 | BD Biosciences | 553347 | |
| PE-Cy5 Rat Anti-Human CD49f | BD Biosciences | 551129 | |
| PE/Cy7 anti-mouse/rat CD29 Antibody | BioLegend | 102222 | |
| Pen/Strep (10,000 U/mL) | Gibco | 15140122 | |
| Rat Tail Collagen I (coating) | Sigma | 122-20 | |
| Rat Tail Collagen I for 3D culture | Cultrex | 3447-020-01 | |
| recombinant human FGF10 | Peprotech | 100-26 | |
| recombinant human Noggin | Peprotech | 120-10C | |
| recombinant murine EGF | Peprotech | 315-09 | |
| recombinant murine FGF basic (bFGF) | Peprotech | 450-33 | |
| RPMI 1640 medium | Gibco | 31870025 | |
| SBTI (Trypsin inhibitor from Glycine max) | Sigma | T6522 | |
| Tris BASE | Roche | 11814273001 | |
| Trypsin-EDTA 0,05% | Gibco | 25300054 | |
| Waymouth medium | Gibco | 31220023 |
参考文献
- Panciera, T. Induction of Expandable Tissue-Specific Stem/Progenitor Cells through Transient Expression of YAP/TAZ. Cell Stem Cell. 19 (6), 725-737 (2016).
- Bar-Nur, O. Lineage conversion induced by pluripotency factors involves transient passage through an iPSC stage. Nat Biotechnol. 33 (7), 761-768 (2015).
- Xu, J., Du, Y., Deng, H. Direct lineage reprogramming: strategies, mechanisms, and applications. Cell Stem Cell. 16 (2), 119-134 (2015).
- Clevers, H. Modeling Development and Disease with Organoids. Cell. 165 (7), 1586-1597 (2016).
- Blanpain, C., Fuchs, E. Stem cell plasticity. Plasticity of epithelial stem cells in tissue regeneration. Science. 344 (6189), 1242281 (2014).
- Bai, H. Yes-associated protein regulates the hepatic response after bile duct ligation. Hepatology. 56 (3), 1097-1107 (2012).
- Cai, J. The Hippo signaling pathway restricts the oncogenic potential of an intestinal regeneration program. Genes Dev. 24 (21), 2383-2388 (2010).
- Elbediwy, A. Integrin signalling regulates YAP and TAZ to control skin homeostasis. Development. 143 (10), 1674-1687 (2016).
- Taniguchi, K. A gp130-Src-YAP module links inflammation to epithelial regeneration. Nature. 519 (7541), 57-62 (2015).
- Zhang, W. Downstream of mutant KRAS, the transcription regulator YAP is essential for neoplastic progression to pancreatic ductal adenocarcinoma. Sci Signal. 7 (324), ra42 (2014).
- Zanconato, F., Cordenonsi, M., Piccolo, S. YAP/TAZ at the Roots of Cancer. Cancer Cell. 29 (6), 783-803 (2016).
- Azzolin, L. YAP/TAZ incorporation in the beta-catenin destruction complex orchestrates the Wnt response. Cell. 158 (1), 157-170 (2014).
- Stingl, J. Purification and unique properties of mammary epithelial stem cells. Nature. 439 (7079), 993-997 (2006).
- Means, A. L. Pancreatic epithelial plasticity mediated by acinar cell transdifferentiation and generation of nestin-positive intermediates. Development. 132 (16), 3767-3776 (2005).
- Shi, G. Maintenance of acinar cell organization is critical to preventing Kras-induced acinar-ductal metaplasia. Oncogene. 32 (15), 1950-1958 (2013).
- Huch, M. Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 32, 2708-2721 (2013).
- Fernandez Vallone, V. Trop2 marks transient gastric fetal epithelium and adult regenerating cells after epithelial damage. Development. 143 (9), 1452-1463 (2016).
- Yanger, K. Robust cellular reprogramming occurs spontaneously during liver regeneration. Genes Dev. 27 (7), 719-724 (2013).
- Yui, S. YAP/TAZ-Dependent Reprogramming of Colonic Epithelium Links ECM Remodeling to Tissue Regeneration. Cell Stem Cell. , (2017).
- Pan, F. C. Spatiotemporal patterns of multipotentiality in Ptf1a-expressing cells during pancreas organogenesis and injury-induced facultative restoration. Development. 140 (4), 751-764 (2013).
- Panciera, T., Azzolin, L., Cordenonsi, M., Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol. 18 (12), 758-770 (2017).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。