
Method Article
Única molécula fluorescência In Situ Hybridization (smFISH) análise em Budding fermento crescimento vegetativo e meiose
Neste Artigo
Resumo
Esta molécula fluorescência em situ da hibridação protocolo é otimizado para quantificar o número de moléculas de RNA em leveduras brotamento durante a meiose e crescimento vegetativo.
Resumo
Única molécula fluorescência situ hibridação in (smFISH) é uma técnica poderosa para estudar a expressão gênica em células únicas devido à sua capacidade de detectar e contar as moléculas de RNA individuais. Complementares aos métodos baseados em sequenciamento profundos, smFISH fornece informações sobre a variação de célula para célula em abundância de transcrição e a Localização subcellular de um determinado RNA. Recentemente, temos usado smFISH para estudar a expressão do gene NDC80 durante a meiose em leveduras brotamento, na qual dois transcrição isoformas existem e a isoforma de transcrição curta tem sua sequência toda compartilhada com o isoform longo. Para identificar com confiança cada isoforma de transcrição, nós otimizadas conhecidos smFISH protocolos e obteve alta consistência e qualidade dos dados de smFISH para as amostras adquiridas durante a brotação fermento meiose. Aqui, descrevemos este protocolo otimizado, os critérios que usamos para determinar se é obtida a alta qualidade dos dados de smFISH e algumas dicas para a aplicação do presente protocolo em outras cepas de leveduras e condições de crescimento.
Introdução
Dinâmica regulação da expressão genética impulsiona o desenvolvimento de um organismo, bem como a sua resposta ao estresse ambiental, infecção e alterações no metabolismo. Estudos que se concentram no Regulamento transcriptional dependem de tecnologias que medem a abundância de RNA. Um tal método, denominado única molécula fluorescência em situ da hibridação (smFISH), é utilizado para detecção de moléculas individuais de RNA em células únicas1,2. Esse método permite que a medição da variabilidade na expressão gênica celular para celular e determinação da localização intracelular de RNA.
A técnica mais comumente usados smFISH, a detecção de uma molécula de RNA individual exige vários curtas sondas de DNA (frequentemente ~ 48 20-mer sondas) que são complementares ao RNA alvo e são conjugadas para a mesma tintura fluorescente. Ligação de uma única sonda fluorescente resulta num sinal fraco, mas o sinal do conjunto de todas as sondas é robusto. Esse recurso melhora a relação sinal-ruído porque mesmo que um único teste pode apresentar vinculação fora do alvo, tal sinal deverá ser muito fraco em relação ao que o destino de molécula de RNA3. No erro de deteção, o número de moléculas de RNA pode ser contado e comparado através de várias condições de crescimento e entre diferentes mutantes.
Desde seu primeiro desenvolvimento, foi adaptada para estudar vários aspectos da expressão gene4, tais como a transcrição alongamento1,2, emenda5,6, transcriptional estourando7 smFISH , 8 , 9, expressão alélica intracelular de11,10,12e RNA localização13,14,15. Recentemente, nós usamos este método para estudar a expressão de duas isoformas de transcrição do gene da mesma, em que a isoforma de transcrição curta (NDC80ORF) tem sua sequência toda compartilhada com o isoform longo (NDC80luti )16. Para identificar com exclusividade as duas isoformas de mRNA, usamos dois conjuntos de sonda: um conjunto é específico para a sequência única de NDC80luti, e o outro conjunto, conjugado com outra tintura fluorescente, vincula-se à região comum das duas isoformas. O NDC80luti RNA é identificado como um ponto colocalized com ambos os sinais fluorescentes, Considerando que o NDC80ORF RNA é aquele que contém apenas o sinal do conjunto de sonda comum. Desde o número da NDC80ORF transcrições é calculado por um método de "subtração", alta eficiência de hibridização da sonda e uma alta relação sinal-ruído são necessários com confiança identificar pontos smFISH e reduzir o erro propagação.
Este artigo descreve um protocolo otimizado para executar smFISH no brotamento levedura Saccharomyces cerevisiae. No presente protocolo, o número do celular, tempo de fixação, tempo de digestão, tampão de digestão, sonda concentração e hibridação buffer usado para os experimentos de smFISH foram otimizados para o SK1 estirpe fundo de S. cerevisiae em fase vegetativo crescimento ou meiose. No entanto, constatamos também neste manuscrito: (1) o método para verificar a qualidade dos dados de smFISH após a aquisição da imagem e (2) os passos no protocolo que podem exigir a otimização adicional para a tensão de diferentes origens e condições de crescimento.
Protocolo
Nota: Todos os buffers e os suportes utilizados neste protocolo são listados na tabela 1. As informações do fornecedor para os reagentes são listadas na Tabela de materiais.
Dia 1/dia 1-2:
Nota: Para a cultura vegetativa, desenvolvem-se células para uma OD600 de 0,4 a 0,6 em meio preferencial. Para cultura meiótica, induzi células submeter-se a meiose, usando um método preferido (normalmente cultivo em uma OD600 de 1,85).
1. fixação e digestão da amostra
- Conserte um total de ~3.5 OD600 de células em formol 3%.
- Para cultura vegetativa, adicione 5,52 mL da cultura a 480 µ l de formol de 37% em tubos cônicos de 15 mL.
- Para cultura meiótica, adicione 1840 µ l de cultura a 160 µ l de formol de 37% em 2 mL microcentrifuge tubos. Inverter ~ 5 vezes para misturar.
Atenção: O formol é tóxico. Manuseie e elimine de acordo com regulamentos institucionais.
- Coloque os tubos em um tambor de cilindro em temperatura ambiente por 20 min. Para amostras meióticas, após a fixação em temperatura ambiente por 20 min, continuam durante a noite de fixação no 4 ˚ c, rotação.
Nota: A fixação durante a noite aumenta a reprodutibilidade e a qualidade dos dados smFISH obtidos para as amostras meióticas. Tempo de fixação deve ser otimizado. - Enquanto as amostras estiver corrigindo, descongele 200mm vanadil Ribonucleoside complexos (VRC) em 65 ˚ c durante pelo menos 10 minutos.
- Preparar a mistura de mestre digestão em um tubo de 15 mL: para 1 amostra, misturar 425 µ l de tampão B com 40 µ l de 200mm VRC (aquecido a 65 ˚ c); para 5 amostras, misture 2125 µ l de tampão B com 200 µ l de 200mm VRC (aquecido a 65 ˚ c). Vórtice ~ 5 s para totalmente resuspenda o VRC antes de adicionar à mistura de mestre, que irá aparecer a luz verde acastanhado após adição da VRC.
Nota: Adição de VRC durante a digestão melhora a consistência dos resultados smFISH, potencialmente, inibindo o contaminante de nuclease introduzido pela mistura de zymolyase, que é purificada do extrato bruto. A quantidade de VRC necessário a esta etapa deve ser otimizada. - Para amostras vegetativas, centrifuga os tubos no ~ 1057 x g durante 3 minutos. Para amostras meióticas (após a fixação da noite), centrifugar a 21.000 x g, durante 1,5 min. decantar ou aspirar o sobrenadante para resíduos de formaldeído.
Nota: Todas as etapas de centrifugação são realizadas à temperatura ambiente. - Ressuspender as células em 1,5 mL de frio Buffer B pipetando para cima e para baixo ou invertendo os tubos e passando rapidamente para misturar. Transferi amostras vegetativas para tubos de 2 mL após ressuspensão.
- Centrifugue a 21.000 x g 1,5 min. remover a maior parte do líquido por aspiração vácuo ou pipetando, deixando para trás ~ 100 µ l.
- Ressuspender as células em 1,5 mL de frio B. Buffer
- Centrifugue a 21.000 x g 1,5 min. remover a maior parte do líquido por aspiração vácuo ou pipetando, deixando para trás ~ 100 µ l.
- Ressuspender as células em 1,5 mL de frio Buffer B. Centrifugue a 21.000 x g 1,5 min. aspirado o líquido completamente por vácuo ou pipetando.
- Ressuspender as células em 425 µ l de mistura de mestre de digestão e brevemente vórtice para ressuspender. Adicione 5 µ l de zymolyase de 10 mg/mL de 100T a cada tubo.
Nota: Vórtice a zymolyase toda vez antes de adicionar aos tubos. Zymolyase adicione a cada tubo individualmente, em vez de adicionar à mistura de mestre. Ambas as etapas ajudam a manter a consistência de digestão entre os tubos, porque zymolyase precipita-se rapidamente. - Vórtice de 2-3 s para misturar. Coloque os tubos em um tambor de cilindro e digerir a 30 ˚ c durante 15-30 min. Para as células vegetativas e primeiros estágios meióticos, normalmente demora ~ 15 min, e para prófase meiótica e divisões meióticas, normalmente demora ~ 30 min.
Nota: Verifique no microscópio cada 5 min depois de 15 min e parar a digestão quando aparecem de ~ 80% das células não-transparente e não-refrativa. De agora em diante, as células são muito frágeis. Lidar com células suavemente e evitar o uso de vácuo aspiração ou num Vortex. - Centrifuga os tubos no ~ 376 x g durante 3 min. Retire o líquido completamente pipetando.
- Resuspenda suavemente células com 1 mL de tampão B pipetando acima e para baixo 1 - 2 vezes para misturar.
- Centrifuga os tubos a ~ 376 x g durante 3 min. líquido de remover o tampão B pipetando. Delicadamente Ressuspender as células em 1 mL de etanol a 70% (diluído com água livre de RNase).
- Incube a temperatura ambiente para 3,5-4 horas.
2. hibridação
- Levar o formamide à temperatura (para alíquota de 50 mL, leva ~ 30 min em banho-maria).
Nota: Não abra a garrafa de formamida até a garrafa atinja a temperatura ambiente para evitar a oxidação de formamida.
Cuidado: Formamida é tóxica. Manuseie e elimine de acordo com regulamentos institucionais. - Prepare o tampão de lavagem de formamida 10% em um tubo cônico de 15 mL.
- Centrifuga os tubos no ~ 376 x g durante 3 min. remover 500 µ l de etanol 70% por pipetagem. Suavemente a pipeta acima e para baixo para Ressuspender as células restantes e então transferir as células para tubos de baixa aderência.
Nota: Utilizando tubos de baixa aderência grandemente reduz a perda de células durante lavagens subsequentes. - Centrifuga os tubos novamente a ~ 376 x g durante 3 min. remover todo o etanol pipetando.
- Adicionar 1 mL de tampão de lavagem de formamida 10% e suavemente pipeta acima e para baixo, 2 - 3 vezes para Ressuspender as células.
- Permitir que as células para sentar-se à temperatura ambiente por ~ 20 min enquanto se prepara a solução de hibridação. Para 1 amostra, misture 50 µ l de tampão de hibridização (sala temp.), 5 µ l de 200mm VRC (aquecido a 65 ˚ c) e 1 µ l de cada sonda (final de 200 nM). Para 5 amostras, misture 250 µ l de tampão de hibridização (sala temp.), 25 µ l de 200mm VRC (aquecido a 65 ˚ c) e 5 µ l de cada sonda (final de 200 nM).
- Descongele o tampão de hibridização (congelado a-20 ˚ c) à temperatura ambiente antes de abrir o tubo para evitar a oxidação de formamida.
- Depois de adicionar a 200mm VRC, vórtice para 5-10 s antes de adicionar as sondas, que são projetados adquiridos comercialmente e reconstituído de acordo com as instruções do fabricante.
- Se duas sondas são incubadas co, adicione 1 µ l da 01:10 diluição do estoque a sonda #1, bem como 1 µ l da 01:10 diluição da unidade populacional de sonda #2, tornar-se uma diluição final de ~ 1: 500 para cada sonda. A concentração necessária para o melhor sinal-para-ruído precisa ser otimizado (ver discussão para detalhes).
- Centrifugar as amostras no ~ 376 x g durante 3 min. Retire o sobrenadante em resíduos perigosos pipetando.
- Adicione pelo menos 50 µ l de solução de hibridização de cada tubo. Agite os tubos para misturar.
- Incube a 30 ˚ c em um tambor de cilindro pelo menos 16 horas no escuro.
Dia 2/dia 3
3. lavar e imagem
- Levar o formamide à temperatura.
- Prepare-se 10% formamida o tampão de lavagem (FWB) em um tubo cônico de 15 mL.
- Retirar os tubos do tambor de rolo e coloque-os em uma caixa de folha-cobertas para proteger da luz.
- Centrifugar as amostras no ~ 376 x g durante 3 min. Retire o sobrenadante em resíduos perigosos pipetando.
- Resuspenda em 1 mL de 10% FWB suavemente pipetando acima e para baixo 2 - 3 vezes.
- Incube a 30 ˚ c por 30 min (não rotativos) na caixa de folha-cobertas.
- Centrifugar as amostras a ~ 376 x g por 3 min. Retire o sobrenadante para resíduos perigosos por pipetagem e deixar ~ 50 µ l.
- Enquanto isso, preparar DAPI/FWB em um tubo cônico de 15 mL: para 1 amostra, misturar 1000 µ l de tampão de lavagem de formamida 10% com 1 µ l de 5mg/mL DAPI. Para 10 amostras, misture 10 mL de tampão de lavagem formamide 10% com 10 µ l de 5mg/mL DAPI. Resuspenda em 1 mL de DAPI/FWB suavemente pipetando acima e para baixo 2 - 3 vezes.
- Incube a 30 ˚ c por 30 min (não rotativos) na caixa de folha-cobertas.
- Descongele os reagentes antilixívia no gelo.
- Centrifugar as amostras no ~ 376 x g durante 3 min. Retire o sobrenadante completamente pipetando. Se necessário, centrifugue novamente para remover todo o sobrenadante.
- Para as amostras que não são imagem imediatamente, resuspenda o pellet em 50 µ l de Buffer GLOX sem enzimas. Pipeta acima e para baixo 3 - 4 vezes para misturar.
- Mantenha todas as amostras unimaged na caixa folha-cobertas no 4 ˚ c até que esteja pronto a imagem. Quando estiver pronto para a imagem, Centrifugar as amostras a ~ 376 x g por 2 min e retire o sobrenadante completamente por pipetagem. Resuspenda em GLOX com enzimas como abaixo.
- Para amostras sendo fotografada imediatamente, adicionar 15 a 20 µ l de tampão GLOX com enzimas. Suavemente a pipeta para cima e para baixo se misturam.
Nota: O volume adicionado pode variar dependendo do tamanho do centrifugado. Recomendamos que resuspending a pelota em 15 µ l de tampão GLOX com enzimas e verificar a densidade de células em microscópio. Se são demasiado densas (células aglutinação uns sobre os outros), adiciona extra reserva GLOX com enzimas. Se as células são muito escassas, centrifugar a amostra e remover ~ 5 µ l de tampão. - Pipetar 5 µ l para uma lamela (18 x 18, n. º 1) e colocar a lamela em um slide.
- Coloca uma limpeza de laboratório em cima do slide onde a lamela é colocada. Pressione suavemente sobre a limpeza de laboratório para definir o slide (deve ver líquido saindo de todas as quatro bordas da lamela).
- Transferi o slide para o quarto de microscópio em uma caixa coberta por papel alumínio.
- Imagem usando um microscópio de fluorescência de campo amplo com alta ampliação (60-100x) e alta abertura numérica.
Observação: Para coletar o número máximo de fótons emitidos por sondas smFISH, microscópios confocal não são recomendados, como eles limitam significativamente a quantidade de luz a serem coletados. Aqui, os dados foram coletados em um microscópio equipado com 100 X, objectivo de at 1.4, usando filtros CY5 (EX632/22, EM679/34) fotografada em 1.3 s, 100% T; TRITC (EX542/27, EM597/45), 1.3 s, 100% T; e DAPI (EX390/18, EM435/48), 0,05-0,1 s, T. 32-50% Uma imagem brilhante-campo referência também deve ser adquirida. Adquirir 15-25 fatias com um tamanho de passo de 0,1-0,2 µm, de inteiramente abaixo o foco do campo de visão inteiramente acima, a fim de garantir que todos os pontos do RNA são contabilizados.
4. análise de imagens
- Analise os dados de smFISH com ferramentas de análise de smFISH publicado como peixe-quant17 e plateia2ou com programas feitos sob medidos, conforme as suas necessidades exatas do experimento.
Nota: Um pipeline de boa análise deve permitir que os usuários determinar um limite para separar os verdadeiros pontos de RNA do fundo e estatísticas, tais como as coordenadas X, Y e Z (opcionais) e a intensidade de cada ponto, o número de pontos em cada célula de saída e possivelmente montagem parâmetros como uma maneira de filtrar as detecções falsas.
Resultados
Para avaliar o quão bem o protocolo smFISH trabalhado (indicado na Figura 1), nós projetamos um conjunto de 54 sondas que o frame de leitura aberto do gene NDC80 de azulejos (Figura 2A, topo). A sonda localizada em mais de 5' extremidade é referida como sonda 1; a próxima vítima, sonda 2; e a terceira, 3 sonda, etc. As 27 sondas atribuído um ímpar número (sonda 1, 3, 5...) são todos conjugados para o corante fluorescente CAL Fluor 590 (CF590); e as 27 sondas atribuído um número par (sonda 2, 4, 6...), conjugado com o corante fluorescente Quasar 670 (Q670). Portanto, esses conjuntos de sonda alternados são muitas vezes referidos como sondas "ímpar". Após a hibridização, ambos os conjuntos de sonda devem rotular a transcrição da mesma.
Após a detecção local, usamos algumas medidas para julgar a qualidade do conjunto de dados smFISH. O primeiro foi o grau e a qualidade de colocalization para as sondas/ímpar. No nosso caso, 88% de todos os pontos de smFISH colocalized (Figura 2A e 2B), com mais de 95% das manchas pareadas dentro de 2 pixels do outro (Figura 2, emparelhado), que está dentro do valor esperado dado qualquer cromática e deteção aberração entre os dois canais fluorescentes. Em comparação, menos de 10% dos pontos da marcação sem paridade tinha uma vizinho mais próximo de distância de menos de 2 pixels, mostrando que a probabilidade de interpretação um par local é baixa (Figura 2). A 12% dos pontos da marcação sem paridade foram divididos uniformemente entre os dois canais (Figura 2B, comparar somente CF590 com somente Q670); e assim, concluímos que este gene 2,4 kb com uma gama de expressão entre zero a 45 cópias por célula, ~ 94% de moléculas de RNA foram detectadas com precisão em cada canal fluorescente. Se o protocolo de smFISH eram suboptimal, um observar (1) uma fração maior de manchas de smFISH com apenas um dos dois sinais fluorescentes (não-colocalized), e/ou (2) células com uma relação sinal / ruído muito baixa ou nenhum sinal em tudo.
Em seguida perguntamos se a deteção do sinal smFISH era inclinada em relação ao número total de moléculas de RNA por célula. Em uma população, o número total de um determinado RNA em cada célula encontra-se em uma distribuição, com algumas células abrigando mais moléculas de RNA do que outros. Um protocolo de boas smFISH robusta deve detectar o RNA independentemente se um número elevado ou baixo número de moléculas de RNA está presente em cada célula. Para testar isso, para cada célula, cálculo da fração de manchas de smFISH com sinais colocalized e a fração com apenas um dos dois sinais fluorescentes. Após o agrupamento das células que tinham o mesmo número de pontos totais por célula, calculamos a fração média de colocalized (combinados) ou não-colocalized (CF590-somente ou Q670) de pontos em um determinado grupo e graficamente esta média em função do número total de manchas por célula (Figura 3). Para cada categoria de pontos, as frações foram semelhantes em toda a gama do número total de pontos por célula. Por exemplo, comparando as células com um total de 20 pontos fluorescentes por célula contra aqueles com um total de 30 pontos, as frações médias dos pontos colocalized foram semelhantes. Este resultado sugere que nosso protocolo poderia detectar moléculas uma gama de RNA em uma célula (até pelo menos 40 ~ moléculas por célula). Se o protocolo funcionou abaixo, um pode observar um viés. Por exemplo, a fração dos pontos com apenas um dos dois sinais fluorescentes pode aumentar como o número total de pontos por célula aumentada.
Usando este protocolo otimizado, examinamos a expressão do gene NDC80 , que codifica uma proteína de cinetócoro, em condições diferentes de crescimento. O gene NDC80 expressa duas isoformas de mRNA: O isoform tempo undecoded transcrição, NDC80luti, tem uma extensão de ~ 400 pares de bases na extremidade 5' em comparação com o curta NDC80ORF isoform, mas ambas as transcrições compartilham região codificante do gene NDC80 (ver esquema na Figura 4A). Nós projetamos dois conjuntos de sondas: conjunto The 590 CF vincula-se à região original 5' de NDC80luti; e o conjunto de 670 Q vincula-se à região comum de NDC80luti e NDC80ORF. As transcrições NDC80luti foram detectadas como as manchas de smFISH onde os sinais de ambos sonda moda colocalized, Considerando que o NDC80ORF transcrições foram aqueles com o sinal apenas de Q 670. Devido ao tamanho do segmento exclusivo de NDC80luti, nós só poderia telha 20 pontas de prova do oligonucleotide ao longo desta região, que é menor do que o recomendado sondas de 30 a 48. Assim, determinamos primeiro se este conjunto de sonda robustamente poderia detectar RNA em nosso protocolo otimizado. Para qualquer canal fluorescente, graficamente a relação sinal-ruído (SNR, definido como a variação de pixels em torno de um ponto em relação a intensidade local) contra o sinal detectado para cada ponto de smFISH e gerado um gráfico de dispersão para os locais de identificado no campo de visão inteiro (imagens de exemplo na Figura 4A ) e parcelas na Figura 4B. Duas populações distintas foram claramente identificadas para ambos o 670 Q e a sonda de 590 CF define (Optimized, Figura 4B), sugerindo que as manchas de smFISH verdadeiro (dentro da área cinza) poderiam ser separadas do sinal de fundo. Observe que, na condição de qualidade inferior, tal separação foi menos óbvio (Suboptimal, Figura 4B).
Usamos esses conjuntos de dois sonda para estudar a expressão da NDC80luti e NDC80ORFdurante o crescimento vegetativo e meiose. Nas células vegetativas, sinal robusto do conjunto sonda 670 Q foi detectado, mas não de 590 a CF sonda conjunto (Figura 5A, vegetativa), concordando com a observação pela mancha do Norte que só o isoform curto foi expressa durante o crescimento vegetative16 . Este resultado também sugere que o conjunto de sonda 590 CF específico, gerando baixo fundo em nosso protocolo otimizado de smFISH. Em contraste, sinal robusto de ambos os conjuntos de sonda foi detectado na prófase meiótica e a maioria dos pontos tinha colocalized de sinal (Figura 5A, prófase meiótica). Juntamente com análise de borrão do Norte (Figura 5B), esta observação confirmou que a isoforma de longa NDC80luti manifestou-se especificamente na meiose. Estes dois tipos de mRNAs foram detectados no citoplasma (fora da região DAPI), sugerindo que ambos foram exportados do núcleo, consistente com o ribossoma, criação de perfil de dados18.
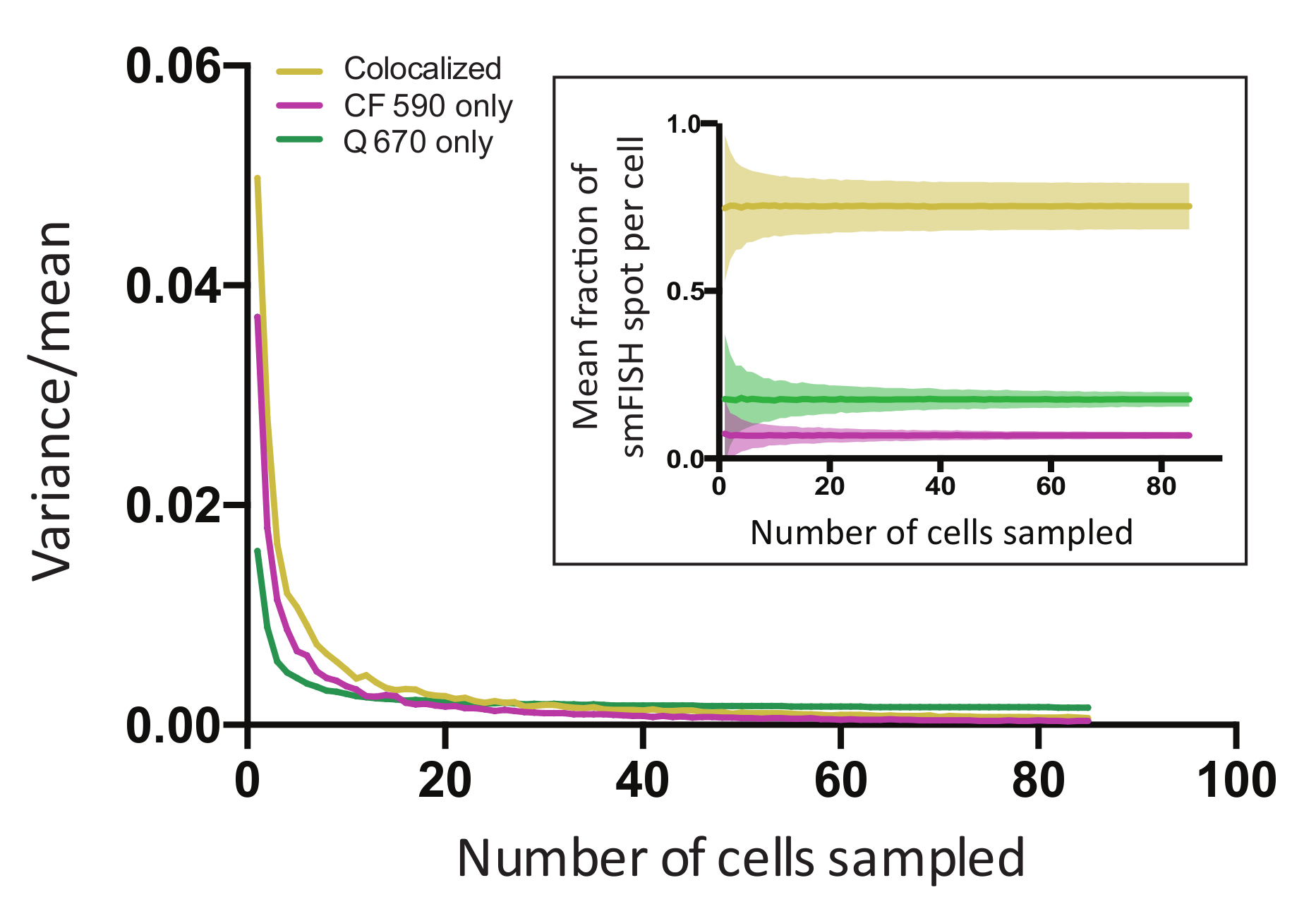
Para determinar se os dados suficientes foram coletados para conta com precisão para a variação biológica intrínseca ao nosso conjunto de dados, realizamos análise de inicialização usando as estatísticas de cada célula, que incluía (1) a fração dos pontos colocalized por célula e (2) a fração das manchas com um dos dois sinais fluorescentes (CF apenas 590 e Q apenas 670). Nesta análise, um programa selecionado aleatoriamente uma célula de 437 células quantificadas para 500 iterações. Em seguida, foram calculados a média e a variância das respectivas estatísticas associadas com esses 500 células selecionadas. Este processo foi repetido então para selecionar aleatoriamente duas células de todas as células, sem substituição; e depois para selecionar aleatoriamente três células, etc. até que uma metade do tamanho do conjunto de dados total foi alcançado. A variância do conjunto de dados se estabilizou após ~ 40 células, sugerindo que, após este ponto a maioria da variação média é intrínseca para os dados ao invés de um artefato de undersampling (Figura 6, inserir). Este efeito se tornou mais evidente quando nós graficamente a variação de amostra, dividida pela média da amostra, em função do número de células amostrados em cada ciclo de iteração (Figura 6). Com os dados mostrados, a mudança na variação tornou-se diminishingly pequena após ~ 60 células. O número de células quantificado em um experimento de smFISH deve exceder o número mínimo de células necessárias para alcançar uma média estável e um platô da variância. No nosso caso, podemos quantificar mais 95 células por amostra, por replicar. O número total de células (> 400) bem ultrapassado o número mínimo de células (~ 60) necessário para refletir a média da população.
Com uma custom-made Matlab programa16, as células vegetativas foram encontradas para ter uma média de 5 NDC80ORF transcrições por célula, enquanto em células da prófase meiótica, a mediana caiu significativamente para 4 transcrições por célula (Wilcoxon bicaudal Teste de soma de Rank, p = 0,026) (Figura 7). O número médio de NDC80luti transcrições por célula foi 21 transcrições, e 100% das células expressa as transcrições NDC80luti . Nós graficamente a fração das células com um determinado número de transcrições como um histograma de frequência relativa, passo a passo, porque o número de moléculas de mRNA é uma quantidade discreta. O maior bin de cada histograma foi normalizada para a mesma altura.

Figura 1: Flow-chart para o protocolo de smFISH. As células são fixos com formaldeído em temperatura ambiente por 20 min ou no 4 ˚ c durante a noite, para amostras de crescimento vegetativo e meióticas amostras, respectivamente. Após a lavagem em tampão B três vezes, as células são digeridas por zymolyase até ~ 70-90% das células são digeridos. As amostras digeridas são então lavadas uma vez com Buffer B para remover o zymolyase e posteriormente permeabilizadas em 70% etanol (EtOH) por ~3.5 horas. Para se preparar para a hibridação, amostras primeiro são incubadas em tampão de lavagem 10% formamida (FWB) por ~ 20 min. Em seguida, as amostras são resuspended em ~ 50 µ l de solução de hibridação, que contém as sondas fluorescentes, para incubação durante a noite, no escuro a 30 ˚ c. Após a hibridização, as amostras são incubadas em FWB 10% por 30 min para lavar as excesso sondas e então incubadas em 10% FWB com 4' 6-diamidino-2-phenylindole (DAPI) para manchar o DNA. Para a amostra fotografada imediatamente após a incubação FWB/DAPI, a amostra é resuspended no buffer GLOX suplementado com catalase, Trolox e glicose oxidase (GLOX + enz); Considerando que, as outras amostras são resuspended no buffer GLOX sem enzimas e armazenadas a 4 ˚ c até ~ 3 horas. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: avaliação da qualidade smFISH usando sondas ímpar/par. (A) top: Diagrama esquemático para os conjuntos de sonda/ímpar. Cinquenta e quatro do oligonucleotide sondas, telhando o gene NDC80 foram projetadas. As sondas ímpares foram rotuladas com um fluoróforo (CF590, mostrado em magenta) e as sondas de pares, com mais um fluoróforo (Q670, mostrado em verde). Fundo: Representante smFISH imagens de células de prófase meiótica adquiriram usando os conjuntos de sonda/ímpar. Foram colhidas amostras 6 horas depois de células (UB8144) foram transferidas para meio de esporulação, uma vez que quando estas células foram presos em prófase meiótica. DNA foi manchado com DAPI (mostrado em azul). Imagens são mostradas como as projeções de intensidade máxima de z-pilhas. Barra de escala: 5 µm. (B) percentagem das manchas de smFISH pareados ou não pareado obtidos usando os conjuntos de sonda/ímpar. Um total de 428 prófase meiótica de células analisadas, pool de dois experimentos independentes. Esta figura é modificada da Figura 2-suplemento Figura 4 de Chen et al . 16 (C) A cumulativa densidade função (CDF) da distância entre cada par de pontos emparelhados e a distância entre o vizinho mais próximo de um local da marcação sem paridade. Para cada ponto detectado em um canal fluorescente, o algoritmo "k mais próximo vizinho" foi aplicado para identificar o local mais próximo de detectado — e a distância até esse ponto — no canal complementar. Localizações que eram mútuas mais próximo vizinhos foram consideradas para ser emparelhado, e uma nova lista foi gerada gravando as detecções emparelhadas, somente CF590 e Q670-somente. Para cada categoria de detecções, um histograma CDF das distâncias foi plotado em Matlab, confirmando que corretamente emparelhados pontos realmente eram muito mais perto na distância do que aqueles sem um correspondente em outro canal local. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: número de fracções de manchas emparelhadas e não emparelhadas, em função do RNA total por célula. Para testar se a detecção e o emparelhamento dos pontos smFISH era inclinado a níveis diferentes de expressão, células individuais foram agrupadas pela expressão de RNA total. As frações médios de detecções emparelhadas, somente CF590 e Q670 somente foram calculadas para cada grupo de células. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Avaliação da qualidade dos dados gerados usando as sondas projetadas para NDC80luti smFISH e NDC80ORF mRNAs. As sondas Q 670 (mostradas em verde) cruzam a região comum compartilhada entre NDC80luti e NDC80ORF mRNAs, Considerando que as sondas de 590 CF (mostradas em magenta) cruzam a região original 5' de NDC80luti . (A) imagens do representante smFISH de NDC80luti e NDC80ORF em prófase meiótica, adquiriu sob otimizada contra condições suboptimal. Cepas UB8144 (otimizado condição) e UB1337 (condição suboptimal) foram induzidos a passam por meiose e fixados durante a prófase meiótica. Essas duas linhagens abrigam sistemas de sincronização diferente para induzir a meiose, mas o uso de qualquer sistema não afeta a qualidade de smFISH (dados não mostrados). Na condição de qualidade inferior, as células foram fixadas em temperatura ambiente por 20 min (sem fixação da noite), digerida com zymolyase sem VRC completada e hibridizado em uma concentração mais baixa da VRC. Imagens são mostradas como as projeções de intensidade máxima de z-pilhas. DNA foi manchado com DAPI (mostrado em azul). Barra de escala: 5 µm. Scatterplots (B) exibindo a relação sinal-ruído (SNR) e o sinal de cada smFISH ponto detectado no campo de visão apresentado na Figura 4A, qualquer canal fluorescente, bem como para as condições otimizadas ou qualidade inferior. Na condição otimizada, duas populações dos smFISH locais estiveram presentes. As manchas de smFISH verdadeiro localizavam-se dentro da área cinza, separa o sinal de fundo. Na condição de qualidade inferior, os mRNAs individuais eram difíceis de distinguir a olho nu, e a separação entre o verdadeiras manchas e fundo depois de executar o software de detecção de ponto foi menos óbvia. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Expressão de NDC80luti e NDC80ORF transcrições são controladas temporalmente. (A) imagens do representante smFISH de NDC80luti e NDC80ORF durante o crescimento vegetativo e meiose. Vegetativas amostras foram tiradas quando células (UB8144) foram crescendo exponencialmente em meio nutritivo rico. 6 horas depois de células (UB8144) foram transferidas para meio de esporulação, uma vez que quando estas células foram presos em prófase meiótica, foram coletadas amostras de prófase meiótica. As sondas Q 670 (mostradas em verde) cruzam a região comum compartilhada entre NDC80luti e NDC80ORF mRNAs, Considerando que as sondas de 590 CF (mostradas em magenta) cruzam a única 5' região de NDC80luti . DNA foi manchado com DAPI (mostrado em azul). Cada célula foi encenada por seu sinal Zip1-GFP, um marcador para prófase meiótica. Nosso protocolo de smFISH preserva o sinal forte do GFP sem modificações adicionais. Crescimento vegetativo: Zip1-GFP negativo. Prófase meiótica: Zip1-GFP positivo. Imagens são mostradas como as projeções de intensidade máxima de z-pilhas. Barra de escala: 5 µm. Esta figura é modificada da Figura 2 de Chen et al . 16. (B) NDC80ORF, NDC80,lutie o nível de proteína (Ndc80p) Ndc80 durante a meiose (UB4074). NDC80luti e NDC80ORF níveis foram determinados pelo Borrão do Norte, e Ndc80p nível foi determinada por immunoblot anti-V5 em pontos de tempo indicado. Hxk1p, carregando controle por immunoblot. Como a tensão UB8144, esta variedade também abriga o GAL4 pGAL-NDT80-ER sincronização sistema19,20. As células foram transferidas para meio de esporulação a 0 hora e libertadas da prisão de paquíteno pela adição de β-estradiol 6 horas mais tarde. A transcrição de NDC80luti robustamente foi detectada na prófase meiótica/S, Considerando que o NDC80ORF transcrições foi predominantemente presentes antes da entrada meiótica (0 hora) e durante as divisões meióticas (7-9 horas). Esta figura é modificada da Figura 6J de Chen et al . 16 Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6: Bootstrap análise realizada para as amostras de prófase meiótica mostradas na Figura 5 . Todas as células quantificadas foram agrupadas, e um determinado número (n) das células foram amostradas aleatoriamente com 500 vezes. A média e o intervalo de confiança de 95% foram calculados para a fração de mRNA emparelhadas e não emparelhadas por célula. Estes dados foram plotados para cada escolha do número n (Inset). Planalto de variância foi visualizado plotando a variância da amostra dividida pela média, em função do número de células (n) amostrados. O número total de células medido (437) grandemente excedido o número no qual o erro se estabilizou (~ 60 células), indicando que os dados adicionais não iria melhorar a confiança nas medições. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7: quantificação dos dados smFISH mostrados na Figura 5 , gráfico como os histogramas de frequência relativa das células com um determinado número de NDC80luti e NDC80ORF transcrições por célula, usando os dados agrupados de três experimentos independentes. Linha tracejada indica o número médio de NDC80luti e NDC80ORF transcrições por célula. Cada histograma foi normalizada para que a altura máxima bin foi o mesmo em todos os histogramas. Um número total de 637 células foram analisado para o crescimento vegetativo e 437 para prófase meiótica. Teste de Wilcoxon Rank Sum bicaudal foi realizada por NDC80luti e NDC80ORF, respectivamente, comparando amostras de prófase meiótica e vegetativo. Esta figura é modificada de Figura 2D de Chen et al . 16 Clique aqui para ver uma versão maior desta figura.
{kind=link}
| Tampão B (estoque de 1 L: 1,2 M sorbitol; tampão de fosfato de potássio 0,1 M, pH 7,5) | 1 x |
| Água livre de nuclease | 500 mL |
| Sorbitol | g 218,6 |
| KH2PO4 | 2,18 g |
| K2HPO4 | g 14,62 |
| Água livre de nuclease | Trazer para o volume final de 1 L |
| * Armazenar no 4 ˚ c em alíquotas de 50 mL após esterilização do filtro | |
| Zymolyase 100T (10 mg/mL) | |
| * Armazenar em-20 ˚ c em alíquotas | |
| Dissolver o pó de zymolyase 10 mg em 1 mL de água MilliQ | |
| E. coli tRNA (10 mg/mL) | |
| * Armazenar em-20 ˚ c em alíquotas | |
| Dissolver o pó de tRNA 10 mg em 1 mL de água MilliQ | |
| etanol a 70% (50 mL) | 1 x (mL) |
| Etanol puro | 35 |
| Água livre de nuclease | 15 |
| * Armazenar à temperatura ambiente | |
| Tampão de hibridização (10ml) | 1 x (mL) |
| Sulfato de dextrano de 50% | 2 |
| Escherichia coli tRNA (10 mg/mL) | 1 |
| 200 mM vanadil ribonucleoside complexo (VRC) | 0.1 |
| BSA, 50 mg/mL | 0,04 |
| 20 x SSC | 1 |
| Formamida | 1 |
| Água livre de nuclease | 4.86 |
| * Armazenar em-20 ˚ c em alíquotas de 250 µ l ou 500 µ l | |
| tampão de lavagem formamide 10% (10ml) | 1 x (mL) |
| Formamida à temperatura ambiente | 1 |
| 20 x SSC | 1 |
| Água livre de nuclease | 8 |
| * fazer fresco, vórtice para 20-30 anos para misturar | |
| solução de glicose 10% | |
| * Armazenar no 4 ˚ c em alíquotas após a esterilização do filtro | |
| Dissolver 1 g de glicose em 10 mL de água livre de nuclease | |
| Glicose oxidase | |
| * Armazenar em-20 ˚ c em alíquotas | |
| Dissolver glicose oxidase de 3,7 mg em 1 mL de 50mm NaOAc, pH 5 | |
| Antilixívia (GLOX) Buffer, sem enzimas (1 mL) | 1 x (µ l) |
| 10% de glicose em água livre de nuclease | 40 |
| 1 M Tris, pH 8.0 | 10 |
| 20 x SSC | 100 |
| Água livre de nuclease | 850 |
| * Certifique-se fresco, também pode armazenar alíquotas no 4 ˚ c | |
| Antilixívia (GLOX) Buffer, com enzimas (50 µ l) | 1 x (µ l) |
| Catalase (vórtice levemente, instala-se facilmente) | 0,5 |
| Glicose oxidase | 0,5 |
| 100mm Trolox (dissolvido em etanol) | 1 |
| Buffer de GLOX | 50 |
| * fazer fresco cada vez, prepara-se em gelo, pode ser armazenado no 4 ˚ c, durante 2-3 horas |
Tabela 1. Buffers e mídia
Discussão
O protocolo aqui apresentado é derivado de outros publicados smFISH protocolos2,3,21,22,23e é especificamente otimizado para o fundo de cepa de levedura SK1. Os parâmetros otimizados incluíam o número do celular, duração de fixação, velocidade de centrifugação e duração, duração da digestão, tampão de digestão, concentração de sonda e tampão de hibridização. Outros parâmetros como temperatura de hibridização e duração não foram otimizados. Nesta seção, nós compartilhamos algumas notas que ajudam a se adaptar a este protocolo para qualquer fundo de cepa de levedura e as condições de crescimento de interesse.
Parede celular de leveduras brotamento é um grande desafio contra a obtenção de imagens de alta qualidade smFISH em brotamento fermento porque a parede celular impede a penetração da sonda. Digestão incompleta da parede celular por zymolyase leva a ineficiente hibridização de sondas e alta variabilidade de célula para célula no sinal. No entanto, digestão excesso pode tornar as células muito frágil, levando a significativa perda de células durante as etapas de lavagem e estourando durante a preparação de slides para a imagem latente da pilha. Assim, otimizando a duração da digestão é crucial. Recomendamos a realização de um experimento piloto smFISH digerindo a mesma amostra para diferentes quantidades de tempo. No nosso caso, obtivemos os dados smFISH melhores se nós paramos a digestão quando tornou-se ~ 80% das células não-refrativa. Normalmente, para a mesma condição de crescimento diferentes cepas mutantes genéticas podem ser digeridas com sincronismo semelhante. No entanto, a duração da digestão diferente quando comparada entre as condições de crescimento diferentes e diferentes estágios da meiose em leveduras brotamento. Uma vez que o calendário é determinado, a qualidade do smFISH é reproduzível. Nota que para cepas mutantes de síntese da parede de celular com defeito e/ou composição, a digestão pode ser concluída em menos de 15 min. utilização de diferentes lotes de zymolyase também ligeiramente pode mudar o tempo de digestão.
Concentração de sonda ideal é necessária para alcançar uma alta relação sinal-ruído. Projetamos e comprou nossas sondas smFISH comercialmente. Boas sondas (geralmente 20-mers) devem ter uma porcentagem de GC variando de 35% a 45%, um espaçamento mínimo de 2 pares de base entre as sondas e baixa reatividade cruzada. Usamos um designer sonda baseado na web do fabricante para gerar uma lista de pontas de prova, e se houvesse suficiente sondas para escolher, nós usaríamos o algoritmo de explosão no banco de dados do genoma de Saccharomyces para eliminar as sondas com mais de 17 pares de base sobrepostos com outras regiões genômicas. Escolhemos a mais fotoestável tintura fluorescente (CAL Fluor 590) para o conjunto de sonda anneals para a região única de NDC80luti (CF. 590), porque neste conjunto, o número de testes que satisfeitos todos os critérios acima (20 testes) foi menor do que o número mínimo recomendado pelo fabricante (~ 25 sondas). Após reconstituir a solução de sonda, seguindo as instruções do fabricante, nós fizemos um 01:10 diluição da solução estoque e armazenados a solução diluída em alíquotas de 5 µ l em-20 ˚ c. Cada alíquota foi usada apenas uma vez. Para a otimização, um deve fazer serial diluições das 01:10 diluído solução e teste a concentração que produz a melhor relação sinal-ruído. Recomendamos fazer uma série de diluição de 1: 250, 1: 500, 1: 1000 e 1: 2000 do estoque original. No nosso caso, um fator de diluição 1: 500 resultou na melhor relação sinal-ruído, para ambos os 590 CF e 670 Q conjuntos de sonda.
Tempo de fixação ideal também é fundamental para o sucesso smFISH em leveduras brotamento. Temos encontrado que a fixação da noite no 4 ˚ c, ao invés de fixação à temperatura ambiente por 20 min, melhorou significativamente a consistência e a qualidade dos resultados smFISH para amostras meióticas. Embora nós não testamos como durante a noite fixação pode impactar amostras vegetativas, fixação à temperatura funcionou bem para esta condição de crescimento. Assim, para otimizar o protocolo smFISH para novas cepas ou condições de crescimento, recomendamos começar com tempo curto fixação à temperatura ambiente e aumentar o tempo de fixação se alta variabilidade do sinal é encontrada.
Em comparação com outros protocolos publicados3,22,23, nosso protocolo usa o inibidor de RNase VRC durante a digestão e uma maior concentração de VRC durante a hibridação. Adição de VRC nessas duas etapas melhorou a consistência dos resultados smFISH, possivelmente por melhor preservar as moléculas do RNA contra a atividade de nuclease, que podem ser introduzidos pela mistura de zymolyase (a enzima é purificada a partir de extratos brutos e pode conter RNase contaminantes). Assim, nós recomendamos usar a quantidade de VRC, conforme listado no nosso protocolo ou mesmo umas concentrações mais elevadas de VRC para otimização.
Uma parcela significativa das células pode ser perdida durante as etapas de lavagem em tampão B e o tampão de lavagem de formamida. Para reduzir a perda de células, pode-se aumentar a velocidade de centrifugação. No nosso caso, usando uma velocidade alta (21.000 x g) para nossas amostras de pelotas durante o Buffer B lava perda celular significativamente reduzida. No entanto, células tornam-se muito frágil após a digestão de zymolyase, portanto não é recomendado alterar a velocidade de centrifugação. Em vez disso, nós sugerimos o uso dos tubos de baixa aderência na Scientific EUA, que ajudam muito as células de pelotas durante as lavagens no tampão de lavagem de formamida. No geral, nosso protocolo pode gerar consistentemente uma monocamada de células densas o bastante para a imagem latente eficiente. Normalmente, 7 campos de vista deve render > 130 células apropriadas para quantificação.
Por último, é importante determinar os parâmetros ideais para e as saídas necessárias de análise de imagem. Para detectar manchas de smFISH, programas de análise publicada comumente filtram as imagens raw usando o kernel gaussiano para remover o sinal de fundo e perguntar aos usuários para determinar a relação sinal-ruído para usar para cada conjunto de imagens de2,17. Infelizmente, não há atualmente um único padrão pelo qual são determinados os parâmetros adequados, e então alguns testes empíricos dessas diferentes configurações é necessário. Para definir os parâmetros necessários para cada um destes passos, um precisa iterativamente diferentes conjuntos de parâmetros de entrada e verificar o quanto os resultados obtidos de cada conjunto corresponde da contagem manual em algumas células representativas. Uma vez que encontra-se um conjunto de parâmetros, pode ser usada para a maioria das imagens obtidas apesar das condições de crescimento diferente e origens genéticas.
Além disso, um pode testar se a projeção de intensidade máxima das imagens smFISH pode ser realizada antes da quantificação22. Este passo simplifica o algoritmo de detecção local e reduz significativamente o tempo de geração de imagens, embora à custa de algumas informações potencialmente úteis sobre pontos individuais, tais como sua localização subcellular. No nosso caso, o número de moléculas de mRNA produzido pelo gene NDC80 foi baixo o suficiente para que as manchas eram bem separadas após esse processamento (não é incomum para as transcrições em brotamento fermento3,7). Em casos onde o colocalization análise é crítico, o pipeline de análise precisa determinar a localização de cada ponto de smFISH em cada canal para avaliar colocalization. Dependendo as perguntas específicas sendo perguntado, outras informações como a intensidade de cada ponto também podem precisar de ser extraído o pipeline para posterior análise. Otimizando a chave entra o protocolo e o pipeline de análise de imagem é crucial na obtenção de alta qualidade de dados smFISH para estudar a questão de interesse.
Divulgações
Os autores não têm nenhum conflito de interesses para divulgar.
Agradecimentos
Agradecemos a Anne Dodson e Stephanie Heinrich para aconselhamento sobre como otimizar o protocolo smFISH, Xavier Darzacq para ajudar com a plataforma de análise, Haiyan Huang para sugestões sobre a análise estatística. Este trabalho foi financiado por fundos da March of Dimes (5-FY15-99), Pew Charitable Trusts (00027344), Damon Runyon Cancer Research Foundation (35-15) e Glenn Foundation para EÜ e um NSF pós-graduação Research Fellowship Grant no. DGE-1106400 de JC.
Materiais
| Name | Company | Catalog Number | Comments |
| BSA, RNase-free (50 mg/mL) | Ambion | AM2616 | Store at -20 °C. |
| Catalase | Sigma | C3515 | Store at 4 °C for short-term. Vortex before use. |
| DAPI | Sigma | D9564 | Store at -20 °C after reconstitution in water. Protect from light. |
| 50% Dextran Sulfate | Milli Pore | S4030 | Store at room temperature. Very viscous liquid. Handle with patience. |
| E. coli tRNA | Sigma | R4251 | Store at -20 °C in aliquots after reconstitution in water. |
| Ethanol (100%, 200 proof) | various | Flammable. | |
| 37% Formaldehyde | Fisher | F79-500 | Store at room temperature. Toxic. Use and dispose with caution. |
| Formamide | Ambion | AM9342 | Store at 4 °C. Open after the temperature equilibrates to room temperature in order to prevent oxidation. Toxic. Use and dispose with caution. |
| Glucose | various | Store at 4 °C after dissolving in nuclease-free water. | |
| Glucose oxidase | Sigma | G2133 | Store at -20 °C in aliquots after reconstitution in 50 mM NaOAc, pH 5. |
| Nuclease-free water | Ambion | AM9932 | Store at room temperature. |
| Potassium phosphate (monobasic and dibasic) | various | Store at room temperature. | |
| Probe library, diluted in TE, pH 8.0 | Biosearch Technologies | Store at -20 °C in aliquots (5 µL) after reconstitution in TE pH 8.0, following the instructions from the manufacturer. Protect from light. | |
| Sorbitol | various | Store at room temperature. | |
| 20X SSC | Ambion | AM9763 | Store at room temperature. |
| TE, pH 8.0 | Ambion | AM9849 | Store at room temperature. |
| 1 M Tris, pH 8.0 | Ambion | AM9856 | Store at room temperature. |
| Trolox | Sigma | 238813 | Store at -20 °C in aliquots. |
| 200 mM Vanadyl ribonucleoside complex (VRC) | NEB | S1402S | Store at -20 °C in aliquots after reconstitution, following the instructions from the manufacturer. |
| Zymolyase 100T | MP Biomedicals | 08320932 | Store at -20 °C in aliquots after reconstitution in water. |
| Low-adhesion tubes | USA Scientific | 1415-2600 | Store at room temperature. |
Referências
- Femino, A. M., Fay, F. S., Fogarty, K., Singer, R. H. Visualization of single RNA transcripts in situ. Science. 280 (5363), 585-590 (1998).
- Raj, A., van den Bogaard, P., Rifkin, S. A., van Oudenaarden, A., Tyagi, S. Imaging individual mRNA molecules using multiple singly labeled probes. Nat Methods. 5 (10), 877-879 (2008).
- Rahman, S., Zenklusen, D. Single-molecule resolution fluorescent in situ hybridization (smFISH) in the yeast S. cerevisiae. Methods Mol Biol. 1042, 33-46 (2013).
- Gaspar, I., Ephrussi, A. Strength in numbers: quantitative single-molecule RNA detection assays. Wiley Interdiscip Rev Dev Biol. 4 (2), 135-150 (2015).
- Vargas, D. Y., et al. Single-molecule imaging of transcriptionally coupled and uncoupled splicing. Cell. 147 (5), 1054-1065 (2011).
- Waks, Z., Klein, A. M., Silver, P. A. Cell-to-cell variability of alternative RNA splicing. Mol Syst Biol. 7, 506(2011).
- Zenklusen, D., Larson, D. R., Singer, R. H. Single-RNA counting reveals alternative modes of gene expression in yeast. Nat Struct Mol Biol. 15 (12), 1263-1271 (2008).
- Raj, A., Peskin, C. S., Tranchina, D., Vargas, D. Y., Tyagi, S. Stochastic mRNA synthesis in mammalian cells. PLoS Biol. 4 (10), e309(2006).
- Senecal, A., et al. Transcription factors modulate c-Fos transcriptional bursts. Cell Rep. 8 (1), 75-83 (2014).
- Levesque, M. J., Ginart, P., Wei, Y., Raj, A. Visualizing SNVs to quantify allele-specific expression in single cells. Nat Methods. 10 (9), 865-867 (2013).
- Hansen, C. H., van Oudenaarden, A. Allele-specific detection of single mRNA molecules in situ. Nat Methods. 10 (9), 869-871 (2013).
- Ginart, P., et al. Visualizing allele-specific expression in single cells reveals epigenetic mosaicism in an H19 loss-of-imprinting mutant. Genes Dev. 30 (5), 567-578 (2016).
- Long, R. M., et al. Mating type switching in yeast controlled by asymmetric localization of ASH1 mRNA. Science. 277 (5324), 383-387 (1997).
- Park, H. Y., Trcek, T., Wells, A. L., Chao, J. A., Singer, R. H. An unbiased analysis method to quantify mRNA localization reveals its correlation with cell motility. Cell Rep. 1 (2), 179-184 (2012).
- Jourdren, L., Delaveau, T., Marquenet, E., Jacq, C., Garcia, M. CORSEN, a new software dedicated to microscope-based 3D distance measurements: mRNA-mitochondria distance, from single-cell to population analyses. RNA. 16 (7), 1301-1307 (2010).
- Chen, J., et al. Kinetochore inactivation by expression of a repressive mRNA. eLife. 6, e27417(2017).
- Mueller, F., et al. FISH-quant: automatic counting of transcripts in 3D FISH images. Nat Methods. 10 (4), 277-278 (2013).
- Brar, G. A., et al. High-resolution view of the yeast meiotic program revealed by ribosome profiling. Science. 335 (6068), 552-557 (2012).
- Carlile, T. M., Amon, A. Meiosis I is established through division-specific translational control of a cyclin. Cell. 133 (2), 280-291 (2008).
- Benjamin, K. R., Zhang, C., Shokat, K. M., Herskowitz, I. Control of landmark events in meiosis by the CDK Cdc28 and the meiosis-specific kinase Ime2. Genes Dev. 17 (12), 1524-1539 (2003).
- Dodson, A. E., Rine, J. Heritable capture of heterochromatin dynamics in Saccharomyces cerevisiae. eLife. 4, e05007(2015).
- Trcek, T., et al. Single-mRNA counting using fluorescent in situ hybridization in budding yeast. Nat Protoc. 7 (2), 408-419 (2012).
- Youk, H., Raj, A., van Oudenaarden, A. Imaging single mRNA molecules in yeast. Methods Enzymol. 470, 429-446 (2010).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados