
Method Article
Um Monitor de Strain Gauge (SGM) para válvula contínua Gape medições em moluscos bivalves em resposta à hipóxia laboratório induzido Diel-ciclismo e pH
Neste Artigo
Resumo
Noções básicas sobre as respostas comportamentais dos bivalves suspensão-alimentadores de variáveis ambientais, tais como oxigênio dissolvido, pode explicar alguns processos do ecossistema. Nós desenvolvemos um monitor de baixo custo, baseado em laboratório, calibre de tensão (SGM) para medir respostas de abertura da válvula de ostra, Crassostrea virginica, hipóxia diel-ciclismo e pH cíclico.
Resumo
Um monitor de gape do calibre de tensão de baixo custo, baseado em laboratório, válvula (SGM) foi desenvolvido para monitorar o comportamento de abertura da válvula de moluscos bivalves em resposta à hipoxia diel-ciclismo. Uma ponte de Wheatstone estava ligada ao extensómetros que foram anexados às conchas de ostras (Crassostrea virginica). Os sinais gravados permitiram a abertura e fechamento dos bivalves para ser gravada continuamente durante dois dias períodos de hipóxia induzida experimentalmente de diel-ciclismo e diel-ciclismo a mudanças no pH. Aqui, descrevemos um protocolo para o desenvolvimento de um monitor de baixo custo Extensômetro e descrever, em um experimento de laboratório de exemplo, como usamos para medir o comportamento de abertura da válvula do Oriental ostras (c. virginica), em resposta à hipoxia diel-ciclismo e mudanças cíclicas no pH. Abertura de válvula foi medida em ostras submetidas a cíclica severa hipoxia (0,6 mg/L) dissolvido condições de oxigênio com e sem mudanças cíclicas no pH, condições cíclicas suave hipóxica (1,7 mg/L) e condições normoxic (7,3 mg/L). Demonstramos que quando deparamos com ostras diel repetidos ciclos, eles rapidamente fechem suas conchas em resposta à hipoxia severa e fechem com um intervalo de tempo à hipoxia suave. Quando normoxia é restaurado, eles rapidamente abrir novamente. As ostras não respondeu às condições de pH cíclico sobrepostas a diel ciclismo hipoxia severa. Em condições de oxigênio reduzido, mais de um terço das ostras fechadas simultaneamente. Demonstramos que ostras respondem à hipoxia diel-andar de bicicleta, que deve ser considerada quando avaliar o comportamento de moluscos bivalves de oxigênio dissolvido. A válvula SGM pode ser usada para avaliar as respostas dos moluscos bivalves para alterações em oxigénio dissolvido ou contaminantes. Técnicas para melhor selo de vedação do válvula gape extensómetros da água do mar precisa de novas melhorias para aumentar a longevidade dos sensores.
Introdução
Hipóxia, ou seja, dissolvido as concentrações de oxigênio [] suficientemente baixo para afetar negativamente biológicas e processos ecológicos, mas muitas vezes funcionalmente definida como [fazer] < 2 mg / L1e anóxia (funcionalmente definida como [fazer] de 0.0-0.2 mg/L) ocorrem mais frequentemente e severamente em águas costeiras do mundo, estuários e o oceano profundo2,3 e são muitas vezes agravada pela crescente eutrofização4,5. Com uma extensão de areal crescente de hipóxia e anóxia, macrofauna são afetados negativamente e perder a medida do habitat e qualidade de habitat. Mudança climática prevê-se a agravar a hipóxia e anóxia6.
Em muitos estuários estratificados, enriquecidos com nutrientes como Chesapeake Bay, Estados Unidos, hipóxia sazonalmente persistente pode prevalecer e pode ocorrer ano após ano2. Além disso, diel-ciclismo de hipóxia é frequente em estuários como a Baía de Chesapeake e outros locais e ocorre tarde durante a noite ou de madrugada no verão7,8.
A maioria dos estudos centraram-se sobre os efeitos da exposição contínua dos organismos para baixo [fazer] e sua tolerância à hipóxia e anóxia9,10,11,12,13,14 . Além disso, estudos têm olhou para a mudança em grande escala na composição de espécies distribuições, abundâncias e espécies em resposta ao prolongado baixa [fazer]4,15. Frequentemente espécies que são muito sensíveis à baixa [,] morrer em massas,16 deslocando as restantes espécies de uma fauna mais jovem, de menor porte, curta duração, como, por exemplo, encontradas no ecossistema Louisiana-Texas prateleira4.
Mudanças comportamentais normalmente precedem comunidade colapso17 e estudos relataram sobre respostas comportamentais dos organismos para estendido baixa [fazer]4,16,17,18,19 ,20,21,22,23,24,25. Estes estudos, no entanto, não se concentre as respostas dos organismos para exposições diel-ciclismo de hipóxia e a natureza flutuante do [fazer] disponibilidade em estuários.
Hipoxia Diel-ciclismo em estuários rasos tem recebido crescente sensibilização como estudos monitorar [fazer] mais frequentemente ao longo dos dias com sondas em estuários16,26. Água pode permanecer hipóxica por horas no final da noite ou de madrugada no verão quando não há nenhuma fotossíntese geradora de oxigênio durante a noite mas o alto consumo de oxigênio de7,de respiração aeróbia16. Também foi encontrado que as marés afetou o diel ciclismo de baixas condições de fazer com a mais extremas mínimos observados quando as marés baixa coincidiu com o fim da noite27. Só depois de várias horas de hipóxia [fazer] voltar a normoxia7,16,28 do ciclo diário.
Para determinar a resposta comportamental de c. virginica hipóxia diel-ciclismo e pH nós monitorados a abertura e fechamento das válvulas de ostras expostas ao laboratório induzido diel cíclico e ciclismo de [fazer] pH. Gape respostas de bivalves têm sido utilizadas para detectar condições ambientais adversas. Tampas de válvula de bivalves em resposta a contaminantes29,30,31, algas tóxicas32,33,34, poluição térmica35,36 , 37,38,39,40, alimentação regime39,41, emersão37,42, fotoperíodo43 diminuir a quantidade de alimentos , 44, pH45,46e combinado de pH e oxigênio dissolvido47 foram medidos. Técnicas de abertura, por exemplo, incluíram observações diretas48,49,13, medições contínuas usando ímãs (Dreissena monitor)50e reed switches ou sensores de fibra óptica 51 que requerem água clara. Além disso, sensores de Hall de força magnética e campo magnético têm sido usados para estudar o mexilhão gape ângulo52,53,54,55e um sistema de indução eletromagnética de alta frequência que pode, a medida da variação da distância entre duas bobinas elétricas que são coladas nas válvulas foi usado56,57,58,59. Uma fonte de alta tensão é necessária para o sistema de indução eletromagnética e poder tem que ser entregue para ambos os lados do escudo52. Este sistema também é comercialmente disponível como o "MOSSELMONITOR" (http://mosselmonitor.nl/).
Em um orçamento apertado pesquisa, construímos um monitor barato Extensômetro (SGM) para medir continuamente gape ostra sobre induzida em laboratório diel ciclismo de [fazer] e pH, em condições de baixa visibilidade. Nosso sistema também é muito mais simples do que os sistemas concorrentes, permitindo que muitos animais para serem instrumentados durante um experimento. Nós queríamos determinar as respostas comportamentais de c. virginica para diel ciclismo severa ([] = 0,6 mg/L) hipóxia com pH de controle (pH = 7,8) e ciclagem de pH (pH = 7,8-7.0), respectivamente e gape respostas a suave ([] = 1,7 mg/L) hipóxia. Além disso, queríamos determinar se as ostras são capazes de responder rapidamente às mudanças em [] sobre o diel ciclo e como eles respondem quando normoxia retorna após um evento hipóxico. Talvez as ostras são otimamente adaptadas ao ambiente rapidamente flutuante que é encontrado em muitos estuários16,27 onde eles vivem. Enquanto mais complexa válvula gape monitores estão disponíveis, a SGM oferece uma técnica de baixo custo que permite medições contínuas de válvula gape nas águas, mesmo em condições de baixa visibilidade.

Figura 1. Ponte de Wheatstone para o aparelho de abertura da válvula. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Os calibre de tensão sensores para monitorar gape bivalves são filmes resistivos em um padrão de Meandro em um revestimento protetor de poliimida. Pequenas quantidades de estirpe modulam a resistência do sensor. Os bivalves flexiona o gage estirpe quando ele gapes causando uma mudança na resistência do sensor. Utilizamos uma ponte de Wheatstone anulação, equilibrada, para cada canal de bivalves, como mostrado na Figura 1 para medir a mudança na resistência do sensor. A ponte de Wheatstone é anulada pelo potenciômetro que permite um ganho bastante elevado ser empregado pelo datalogger. Uma ponte de Wheatstone é um método padrão para medir com precisão uma resistência desconhecida usando um rácio para uma resistência conhecida padrão e um voltímetro. A história desta técnica muito antiga é discutida em Ekelof (2001)60. 12 canais, cada um com sua própria ponte de Wheatstone e anulação potenciómetro, integrados a unidade Strain Gauge Monitor (SGM).
Protocolo
1. construção da ponte de Wheatstone para o aparelho de Gape de válvula
Nota: O calibre de tensão é nominalmente 1000 Ω, tão totalmente equilibrar e nulo a ponte, todos os componentes devem ser Ω de 1000.
- Como na Figura 1, solda kΩ dois 1 resistores de precisão uns aos outros e depois para um resistor Ω ~ 976 e um Ω 100 10 Gire o potenciômetro. O intervalo de calibre de tensão típica é Ω alguns fora a 1000 Ω nominal, tão somente que essa resistência precisa ser correspondido. Use um resistor de 976 Ω fixo e um resistor ajustável (10 girar o Poti) para essa finalidade.
- Trazer um lado do braço variável e um lado do kΩ 1 resistor para um conector para anexar o Extensômetro bivalves como mostrado na Figura 1.
- Anexe a cima e em baixo da ponte para uma fonte de alimentação para fornecer a polarização atual para a ponte, como mostrado na Figura 1.
- Prenda os pontos médios da ponte o pré-amplificador e o gravador de dados, como mostrado na Figura 1. Isto irá gravar a resposta bivalves como seu movimento gape modula a resistência do calibre de tensão.
- Depois que o Extensômetro bivalves é anexado, nula a ponte ajustando a 10 vire potenciômetro até que a tensão na entrada do gravador de dados é zero. Use um separado conector BNC e uma mão realizaram voltímetro para essa finalidade.
Nota: Anula a saída significa que ganhos muito altos podem ser usados com o gravador de dados sem saturar a entrada, permitindo tensões muito precisas (gape resposta) a ser medido.
Nota: Nossa configuração multicanal usa mão de componentes discretos, soldada em uma parte padrão de eletrônica "perf-placa". Isso poderia facilmente ser formalizado em uma placa de circuito muito barata com pouco esforço adicional.
2. geral configuração para pH hipóxia Diel-ciclismo e cíclico
- Manter as ostras com conhecida doença de baixa prevalência61 em aquários de 75-L sob uma luz de 14 h: fotoperíodo escuro de 10 h, 7 dias por semana48e expor-lhes alterações cíclicas em [fazer] imitando diel-ciclismo hipóxia como encontrados na natureza7, 8. durante 14 h de luz, simular os níveis claros em um dia ensolarado no 2-m de profundidade em Rhode River, Maryland61.
- Configurar a hipóxia diel-ciclismo e pH cíclico no laboratório para experiências a longo prazo, como descrito em detalhe por Burrell et al (2015) 49. Manipulate [fazer] e pH em cada aquário e se apegam a níveis pré-determinados em todos os momentos. Para evitar o fechamento da válvula devido a limitação de alimento, adicione fitoplâncton62 para os aquários. Compensa qualquer captação potencial da decomposição das algas regulando [fazer] em cada aquário.
3. bivalves Acclimation
- ACCLIMATE ostras à hipoxia diel-ciclismo para cerca de uma semana antes de realizar experimentos gape.
4. criação de fases de platô de hipóxia Diel-ciclismo

Figura 2 . [Fazer] foi manipulada, experimentalmente, todos os dias para gerar hipóxia ciclismo diel. Aqui, 2 ciclos consecutivos diel são mostrados sobre qual gape foram efectuadas medições. Cada ciclo diel contém quatro fases de [fazer]: um = supersaturação (SS), b = normoxia depois supersaturação, c = baixo platô (LP), d = normoxia depois baixo platô. Para o planalto de baixa, hipoxia severa ([] = 0,6 mg/L) com controle ou ciclagem de pH é indicada com uma linha preta grossa, hipóxia leve ([] = 1,7 mg/L) com o ciclismo pH com uma fina linha cinza e normoxia ([] = 7,3 mg/L) com uma linha tracejada. Caixas cinzentas indicam 10-h intervalos escuros durante dois dias. Clique aqui para ver uma versão maior desta figura.
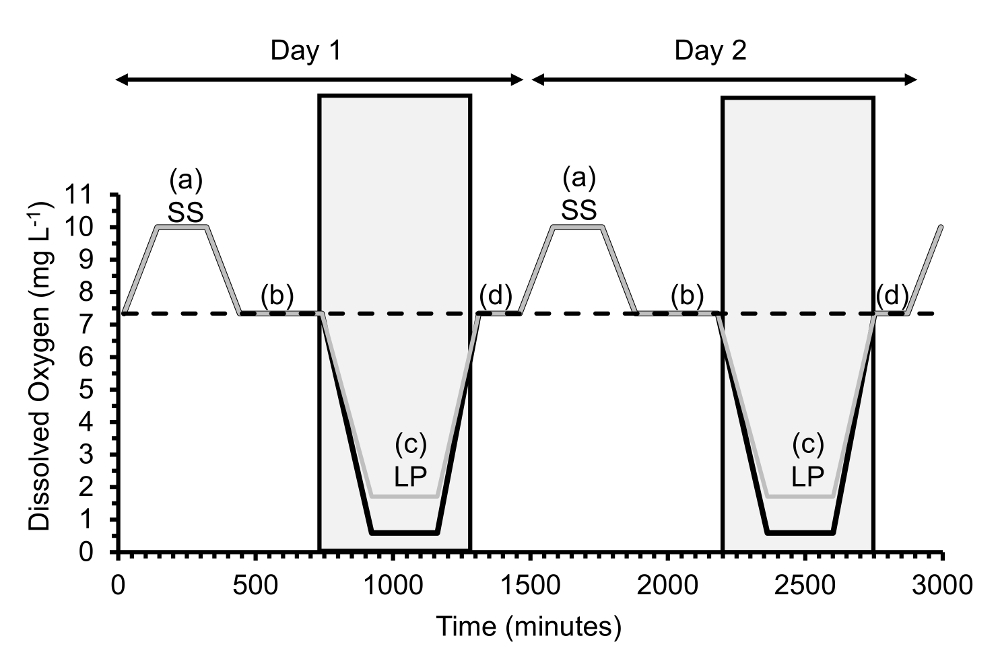{kind=link}
- Para criar o ciclismo hipóxia diel, ajustar [fazer] ao longo de um dia em quatro fases consecutivas onde [fazer] é mantida constante: supersaturação de 2 h, 6 h normoxia depois supersaturação, planalto de baixa 4h (normoxia no caso os controles) e normoxia 2h após a baixa fases do planalto.
- Durante a fase de platô baixo, manter o tratamento normoxic constante em um [fazer] de 7,3 mg/L, a constante tratamento levemente hipóxica em um [fazer] de 1,7 mg/L e a constante de tratamentos de hipoxia severa em um [fazer] de 0,6 mg/L.
- Para hipoxia severa, execute dois tratamentos de pH durante o platô baixo, ou seja, grave hipóxia com pH de controle no qual o pH é mantido em 7,8 e hipoxia severa com pH cíclico onde o pH é um ciclo de 7,8 para 7.0 durante o platô baixo. Execute o tratamento levemente hipóxico com pH cíclico onde pH é um ciclo de 7,8 a 7.0. Execute um segundo tratamento levemente hipóxico com pH constante, se desejado.
5. ramping acima/abaixo de [fazer] entre os estágios de planalto
- Agendar a rampa acima ou abaixo de [fazer] entre as quatro fases constantes para levar 2 h cada, exceto para a rampa abaixo de normoxia depois supersaturação à fase de platô baixo, que leva 3 h. frequentemente, hipóxia foi encontrada para ocorrer tarde da noite ou no início da manhã na Verão de7,8; assim, manter a fase de platô baixo no escuro, bem como os períodos de rampa abaixo ou acima dele, para um comprimento total de escuridão de 10 h.
- Repita a hipóxia diel-ciclismo 4-5 dias uma semana61.
- Adicione as ostras com sensores na fase de arranque antes de supersaturação e removê-los, depois fechá-las, dois dias mais tarde assim que experimentam dois ciclos completos diel (Figura 2) com sensores ligados.
6. fazendo o cabo do Sensor

Figura 3 . Sensor de válvula bivalves SGM Extensômetro. (um) sensor Strain gauge, calibre de tensão (b) anexado a uma ostra mostrando um loop da estirpe de bitola entre ambas as válvulas, sensor de calibre de tensão (c) montado em uma ostra. Clique aqui para ver uma versão maior desta figura.
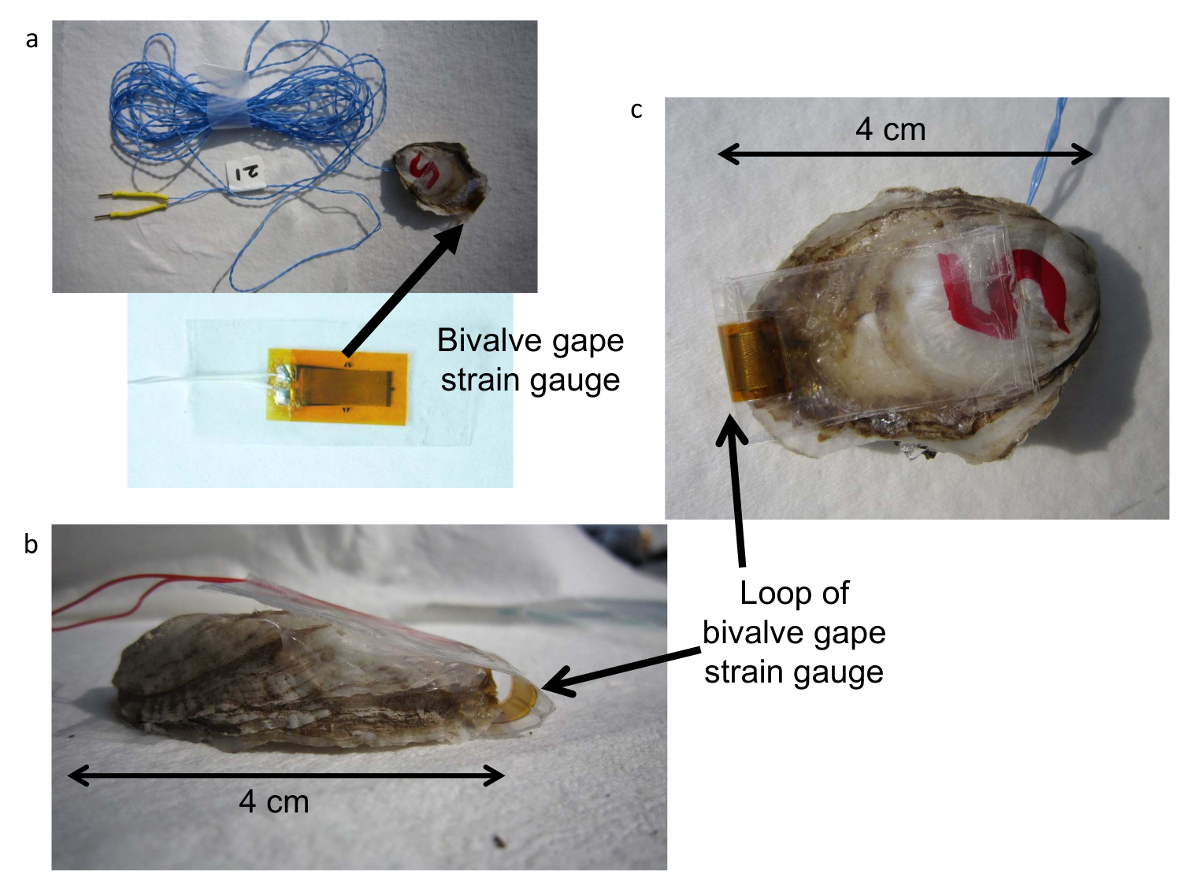{kind=link}
- Construção de cabos de sensor de abertura de válvula rodando dois 32 AWG (7/40) fios (ver Tabela de materiais) da desejada cor (Figura 3a). Corte um comprimento de fio duas vezes o comprimento do cabo do sensor gape desejada, ou seja, duas vezes o comprimento do aquário para a SGM, mais um extra.
- Tem duas pessoas ficar frente a frente, cada um segurando uma extremidade do cabo.
- Deslize a alça de um par de tesouras sobre o cabo de modo que no meio do cabo, um par de tesouras é realizado pelo cabo. Mantenha o cabo sob tensão em todos os momentos (não pode ficar frouxo).
- Têm ambos os povos torcer o cabo na direção oposta, mantendo o cabo sob tensão. Como o cabo começa a torcer, penetrará o par de tesouras no meio (que fornece o peso), e ambas as extremidades do cabo podem girar em torno de si.
- Juntar as duas extremidades do cabo e seguram alto, não permitindo a tesoura com o cabo para tocar o chão ou ir frouxo. Depois acabou-se o cabo de torção, corte o cabo do outro lado há 4 cabo leva total, dois em cada extremidade.
- Como alternativa, para acelerar esta etapa, ter uma pessoa segura uma broca com os fios presos em uma extremidade, manter a tensão nos fios e torcer os fios ao ligar a furadeira.
- Faixa de 2 cm de isolamento de fios em cada extremidade do par trançado.
7. o Sensor de solda
- Estanho o cabo exposto.
- Solda pinos macho crimp Dsub sobre os dois terminais em uma extremidade do cabo do sensor pretinning os pinos e inserindo os cabos.
- Então, isole os condutores colocando psiquiatra tubo ao longo de cada pista. Escolha o psiquiatra tubulação que não é muito maior em diâmetro que as ligações do cabo. Encolha encolher o calor com uma pistola de calor. Tubulação da psiquiatra deve agora ser firmemente enrolada os contatos.
Nota: Estes dois cabos começam plugged na ponte de Wheatstone durante as experiências. - Solda de extremidade restante do cabo sensor para os dois contatos de uma estirpe de 1000 Ω calibre com dimensões de grades de 13,5 mm * 5,5 mm sobre os restantes dois leva (Figura 3a).
- Adicione um sensor para identificar a marca para o cabo do sensor (Figura 3a).
8. o Sensor de vedação
Nota: Os extensómetros corroer rapidamente em água salgada e, portanto, deve ser muito bem lacrados. A qualidade do selo afeta a vida útil do sensor.
- Sele os extensómetros, anexando-os a PCT-2A fita deixando 15 mm de fita nas extremidades longas e 2mm nas laterais do sensor (Figura 3a). Depois de anexar o PCT-2A fita na parte inferior do calibre de tensão, adicione gel de rápida sobre os contatos do sensor e as anexado extremidades abertas dos cabos.
- Anexar um segundo pedaço de fita de PCT-2A do mesmo tamanho como a parte inferior para o lado superior do sensor e unir os dois pedaços de fita com o calibre de tensão no meio junto. Não permita que o gel rápido ao fluxo de outros em qualquer lugar do que sobre os fios e os cabos do sensor.
- Para melhorar a vedação, adicione tiras finas de PCT-2A fita nas bordas onde se encontram os dois pedaços de fita em torno do sensor. Faça isso por três bordas expostas dos sensores.
- Repita as etapas de 6.1 a 8.4 onze vezes para construir 12 cabos de sensor com sensores, um para cada canal na ponte de Wheatstone.
Nota: Bem sucedida selagem dos extensómetros era a variável-chave que afetam a sobrevivência de sensor de bivalves gape em água salgada e para minimizar o ocasionais "saltos" na saída do sensor. Ensaios anteriores sem vedação técnica descrita acima resultaram em corrosão precoce do sensor e subsequente falha ou tração rápida além do intervalo de medição. Melhorias futuras devem incluir técnicas para melhorar a vedação.
9. fixar o Sensor do Gape válvula um Bivalve (Figura 3b, c)
- Remover a ostra de diel-ciclismo aquário hipóxia antes da fase de platô baixo.
- Com esmalte escreva um número de identificação no escudo de cada c. virginicalimpo e seco.
- Anexe os extensómetros selados para as válvulas limpas e secas ao vivo c. virginica (Figura 3b, c) usando o vedador de aquário para permitir que o sensor deve ser removido após a execução. Conecte o sensor de abertura com os contatos da válvula direita (plano) de ostra meio no reservatório (Figura 3c) e deixe um loop do calibre de tensão (Figura 3b) do outro lado as válvulas para não impedir a abertura das válvulas ( Figura 3b). Cole a extremidade livre restante do sensor da válvula à esquerda.
Nota: As observações confirmam que as ostras abrir suas válvulas normalmente com os sensores no lugar. - Não importa qual caminho à volta do sensor é colado, no entanto, a direção do sinal será alterada se algumas pistas estão coladas a válvula à esquerda e alguns na válvula da direita. A direção do sinal torna-se claro o encerramento forçado dos bivalves no final de uma corrida. Para todos os bivalves, colando a parte do calibre de tensão com o sensor leva para o mesmo lado da válvula para uma direção consistente da mudança no sinal de resposta do gape.
- Durante cada etapa do processo de selagem sensores e anexá-las para as ostras, verifique a resistência do sensor periodicamente para certificar-se de que ele permanece dentro do intervalo da eletrônica de leitura e para verificar se há danos potenciais do sensor durante o processamento.
- Anexe 12 ostras 12 sensores para preencher todos os 12 canais do SGM.
- Adicione as ostras para os aquários diferentes apenas antes da fase de platô de supersaturação.
10. instalação de Datalogger e multiplexador para o aparelho de Gape de válvula
- Anexar um sensor para os bivalves e em seguida conecte o cabo para a SGM com pontes de Wheatstone individuais (Figura 1) para cada sensor (um total de 12 pontes) e conectado a um datalogger.
- Definir o data logger período de amostragem para ler uma vez cada 2,5 s e em seguida em média por 15 s.
Nota: A memória de amostra k 62 sobre o datalogger usado aqui permitidos 12 h de operação autônoma. Dataloggers maiores e taxas de amostragem mais rápidas podem ser selecionadas, se desejado. - O sinal de abertura para cada sensor individual de zero após fixar o sensor um bivalve usando o potenciômetro disca sobre a SGM.
- Quando necessário, como em salinidades superiores, corrigir cada sensor periodicamente para drift na resposta do sensor para que o sensor permanece na faixa da SGM (-25 a + 25 mV) saída da ponte de Wheatstone. A gama SGM foi escolhida para sensibilidade máxima entre a ponte de Wheatstone e a entrada do datalogger.
11. implantação de Bivalves com sensores Gape em hipóxia Diel-ciclismo
- Após o aquário selante curou (ca 30 min) retornar os bivalves para um balde de exploração oxigenado com escoamento de água pelo menos 5 h permitir que o selante de aquário lixiviar antes submersão no experimento.
- Coloque as ostras nos aquários durante a fase de arranque para o planalto de supersaturação (SS) (Figura 2).
12. recuperação de Bivalves com sensores Gape
- Ao recuperar a ostra do aquário com diel-ciclismo hypoxiaat o fim do prazo de dois dias, use os dedos para acionar a ostra para fechar e produzir um sinal de fechamento de tensão sensor. Desta forma, a direção e a magnitude do sinal de fechamento podem ser gravados
- Defina a ostra como sistema aberto, se o valor do sensor é maior do que ½ do valor fechado. Defina a ostra como fechado se o valor do sensor é menor do que ½ o valor de fechamento.
Nota: Ostras orientais não abrir muito na faixa intermediária.
13. linearização de válvula cru Gape dados

Figura 4. Linearização dos dados brutos gape de uma ostra. (a) luz 14 h: fases escuras de 10 h, (b) [DO] por um período de dois dias com períodos de hipoxia severa ([fazem] 0,6 mg/L), (c) dados Raw gape tempo série de uma ostra. Linear deriva dos dados é causada pela corrosão do sensor e tem que ser removida no pós-processamento, (d) gape séries temporais de dados depois de pós-processamento, (e) tempo aproximado do real. Para determinar a magnitude e a direção de spike o fechamento, a ostra é acionada para fechar no final de uma corrida. Magnitudes variam entre diferentes sensores. Clique aqui para ver uma versão maior desta figura.
{kind=link}
- Visualmente identificar atividade do gape de ostra e usar o fechado (ou abrir, qualquer um é mais longo) fases dos bivalves para determinar deriva do instrumento. Usar instruções IF no software de planilha para marcar os valores fechados com um e abrir valores com zero (por exemplo, = se (> 5, 1 (verdadeiro), 0 (falso) de células)).
- Remova as fases mais curtas e execute uma regressão linear sobre os restantes dados brutos. Use a equação de regressão para linearizar os dados experimentais (Figura 4). Linearização tem que ser feito para cada sensor individualmente.
Nota: Deriva do sensor é linear através do tempo (dias) tempo e curta (minutos) escalas e a quantidade de tração varia de sensor para sensor. Drift é normalmente negligenciável sobre vãos de curto período de tempo como uma hora, em que apenas ocasionalmente tração linear tem sido constatada. Deriva de sensor exige que calibrações de abertura da válvula e gape sensor tem que ser feito para cada sensor separadamente e que uma linearização tem que ser feito para cada sensor em pós-processamento dos dados.
14. calibração de sensores: largura real Gape contra Sensor mV
Em um experimento de lado, tensão do sensor pode ser calibrado para a largura real gape de ostras.
- Em um aquário, configurar uma ostra com um sensor de abertura. Inicie gravação gape. Acionar a ostra para fechar por tocá-lo. Logo depois, a ostra se abrirá novamente.
- Como a ostra está abrindo, tirar fotografias e anote a tensão do sensor do aparelho gape cada vez que uma fotografia é tirada. Manter o controle de qual foto vai com qual gape a leitura de aparelhos. Coloque uma escala de referência na fotografia, no mesmo nível das válvulas.
- Analisar a distância a ostra é aberta em cada fotografia (mm) usando um programa de processamento de imagem de código aberto como o ImageJ.
- Compare a largura real gape a tensão do sensor e fazer uma análise de regressão. Isto dá a calibração para o status de abertura real e ainda mais as medições do sensor então podem ser relacionadas a largura real gape dos bivalves.
Nota: Enquanto isso mostra gape largura linearmente está relacionada à tensão do sensor, na verdade, este processo demora muito e não foi realizado para os experimentos de gape de hipóxia. Para os experimentos de gape hipóxia foi importante remover as ostras para a menor quantidade de tempo necessário de seus aquários para equipá-los com sensores e devolvê-los para os aquários. Para alguns experimentos, a medição de largura de abertura pode ser importante e as técnicas de calibração acima devem ser seguidas.
Resultados
Ostras expostas a normoxic ininterrupto de água estuarinas (sem hipóxia durante a fase de platô baixa do ciclo diário) estavam abertas a maior parte do tempo e apenas brevemente fechado com pouca frequência (Figura 5). Quando eles fecharam variadas da ostra de ostra. Esse padrão também foi encontrado por Loosanoff e Nomejko 194644 e Higgins 198039. Ostras também não respondeu à diferença nas fases claras e escuras.

Figura 5 . (a-g) Um subconjunto das respostas gape de sete ostras expostas a (h) dois dias de normoxia ([] = 7,3 mg/L) ocasionalmente, feche cada ostra com um padrão diferente. Nighttime simulado é indicado em barras pretas, simuladas dia hora com barras brancas. Esta figura foi modificada de Porter e Breitburg 201663. Clique aqui para ver uma versão maior desta figura.
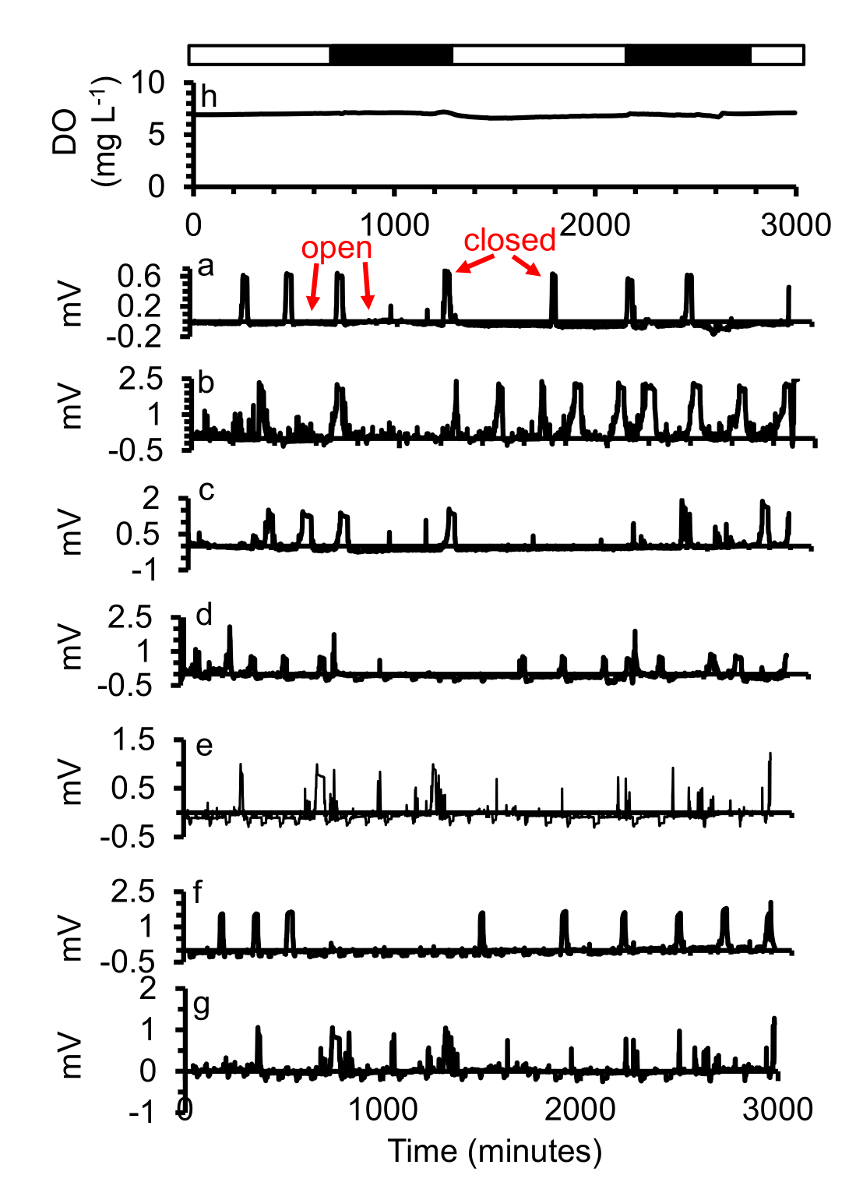{kind=link}
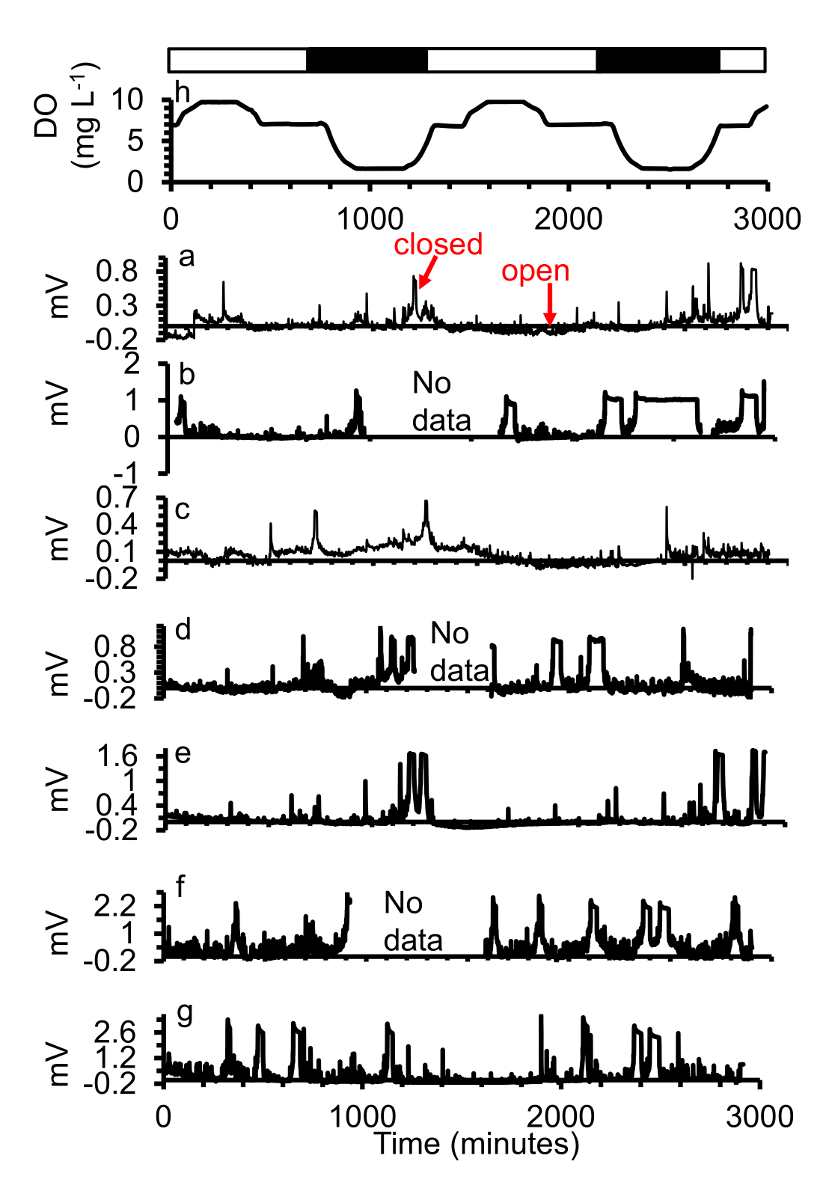
A maioria das ostras expostas à hipoxia severa ([] = 0,6 mg/L) durante o platô baixo fase durante hipóxia diel-ciclismo fechada logo depois o alvo [fazer] foi alcançado, e umas ostras mesmo fechadas antes que o alvo [fazer] foi atingido e permaneceu fecharam durante a maior parte do tempo que hipoxia severa continuou (Figura 6). Controle e pH cíclica não afetou gape.

Figura 6. (a-g) Um subconjunto das respostas gape de sete ostras expostas a (h) dois dias de hipóxia diel-ciclismo com hipoxia severa ([] = 0,6 mg/L) durante a fase de platô baixo. Nighttime simulado é indicado em barras pretas, simuladas dia hora com barras brancas. Esta figura foi modificada de Porter e Breitburg 201663. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Com hipóxia leve ([] = 1,7 mg/L), fechamentos, muitas vezes ocorreram mais tarde durante a fase de platô baixo em vez de no ponto quando o alvo [fazer] foi alcançado (Figura 7).

Figura 7. (a-g) Um subconjunto das respostas gape de sete ostras expostas a (h) dois dias de hipóxia diel-ciclismo com hipóxia leve ([] = 1,7 mg/L) durante a fase de platô baixo. Nighttime simulado é indicado em barras pretas, simuladas dia hora com barras brancas. Energia foi interrompida em c, e, g ("sem dados"). Esta figura foi modificada de Porter e Breitburg 201663. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Durante a fase de platô baixo com variadas [fazer], ostras sob severa ([] = 0,6 mg/L) diel-ciclismo hipóxia com pH de controle (pH = 7,8) bem como cíclico pH (pH = 7,8–7.0) fecharam-se significativamente mais do que as ostras sob normoxia (7,3 mg / L). Lá não houve diferenças significativas na quantidade de encerramento entre o controle e cíclico pH sob hipoxia severa. Ostras sob suave ([] = 1,7 mg/L) diel-ciclismo hipóxia foram fechados quantidades semelhantes de tempo durante o platô baixo como ostras sob hipoxia severa, com controle de pH ou pH cíclico. Além disso, ostras sob hipóxia leve diel-ciclismo fecharam-se uma quantidade similar de tempo como as ostras sob normoxia durante a fase de platô baixo. Lá não houve diferenças significativas na quantidade de encerramento entre o controle e cíclico pH sob hipoxia severa em qualquer das fases.
Durante a fase de normoxia após o baixo platô, ostras que experimentou hipoxia severa durante o platô baixo estavam abertas a maior parte do tempo (Figura 8) e muitas vezes abriram enquanto níveis [fazer] foram aumentando, mesmo antes do planalto de normoxia tinha sido alcançado ( A Figura 6). Durante a supersaturação e a normoxia após as fases de supersaturação, respectivamente, as ostras de todos os tratamentos foram abertas na maioria das vezes. Ostras mostraram que esses padrões sobre diel repetido ciclos ao longo de dois dias de tempo.

Figura 8. Por cento tempo ostras foram fechadas durante o Planalto (um) baixo, (b) normoxia após o baixo platô, supersaturação (c) e (d) normoxia após suspersaturation fases durante o diel ciclismo de [fazer]. Durante a fase de platô baixo em (a): "0.6, cíclico" = [fazer] de 0,6 mg/L (hipoxia severa), cíclico pH (pH = 7,8–7.0) (n = 4); "controle de 0,6," = [fazer] de 0,6 mg/L (hipoxia severa), controlar o pH (pH = 7,8) (n = 4); "1.7, cíclico" = [fazer] de 1,7 mg/L (hipóxia leve), cíclico pH (pH = 7,8–7.0) (n = 4); "controle, controlar" = [fazer] de 7,3 mg (normoxia), controlar o pH (pH = 7,8) (n = 4). Diferenças estatísticas são indicadas por letras diferentes acima das barras (aninhados ANOVA, p≤ 0,05). Barras de erro são erros-padrão da média. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Durante a fase de platô baixo, 1/3 ou mais ostras de hipóxia severa, pH cíclica foram fechadas 70% do tempo e as ostras de hipóxia severa, controlam pH foram fechadas 49% do tempo (Figura 9). Em contraste, durante a fase de platô baixo, 1/3 ou mais ostras do pH cíclico hipóxica suave foram fechado 29% do tempo e do normoxic controle pH tratamento 12% do tempo. As maioria das ostras que fecharam simultaneamente durante a fase de platô baixo, foram de 82% e 67% das ostras na hipóxia severa, pH cíclico e hipóxia grave, controlam tratamentos de pH, respectivamente. De pH hipóxico, cíclico suave tratamento, um máximo de 45% das ostras fechadas durante o platô baixo e ostras expostos a apenas normoxia um máximo de 42% fechado brevemente simultaneamente. Na normoxia após o baixo platô, a supersaturação e a normoxia depois supersaturação, raramente mais do que 1/3 de ostras fechada a qualquer momento.

Figura 9 . Por cento de ostras fechado ao mesmo tempo durante o baixo platô, normoxia, depois baixo platô, supersaturação e normoxia depois fases supersaturação retratado emFigura 2. Para (a), o baixo platô foi severamente hipóxico ([] = 0,6 mg/L) e tinha cíclico pH (pH = 7,8–7.0) (n = 13 ostras). Para (b), o planalto de baixo foi ligeiramente hipóxico ([] = 1,7 mg/L) e tinha cíclico pH (pH = 7,8–7.0) (n = 11 ostras). Para (c), o baixo platô foi severamente hipóxico ([] = 0,6 mg/L) e tinha o controle de pH (pH = 7,8) (n = 11 ostras). Para (d), a fase de platô baixo foi normoxic ([] = 7,3 mg/L) (n = 11 ostras). Linha horizontal retrata onde um terço das ostras foram fechados simultaneamente. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Gape largura está linearmente relacionada com sensor mV (Figura 10). A direção que o sensor é colado na válvula determina a direção do sinal.

Figura 10 . Calibrações de tensão do sensor gape gape real largura do ostras. (a, b) Sensor de abertura leva aposta para a válvula certa das ostras, sensor de abertura (c, d) leva aposta para a válvula à esquerda. Para obter consistência na direção da resposta, os sensores devem ser apostos na mesma direção da válvula de cada ostra. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Típico de estudos enfocam o contínuo, estendido períodos de tempo, das condições de baixo oxigênio e a resposta, muitas vezes medido como a sobrevivência, dos animais. No entanto, neste momento, nosso entendimento das respostas comportamentais dos animais à hipoxia diel-ciclismo é mínima63. Assim, mais estudos devem centrar-se sobre o comportamento dos organismos em resposta à hipoxia diel-ciclismo que ocorre regularmente durante o verão em muitos estuários7,8.
Aqui, apresentamos um método de medir continuamente as respostas comportamentais de moluscos bivalves hipóxia diel-ciclismo e pH cíclico. As ostras não respondeu comportamentalmente para o diel-ciclagem de pH. No entanto, ostras responderam fortemente a condições de baixo oxigênio reduzindo gape de concha. Além disso, mais de um terço das ostras fechadas simultaneamente por longos períodos de tempo sob os tratamentos hipóxicos mas mais abriu novamente sob a fase normoxic que se seguiram. Aumentando o gape e reduzindo o fechamento da válvula para compensar estendido vezes da válvula de fechamento durante a hipoxia severa poderia reduzir os efeitos da hipóxia diel-ciclismo nas ostras se e o fitoplâncton filtram. Assim, tal compensação comportamental poderia minimizar web comida ou efeitos do ecossistema.
Um passo fundamental neste método de SGM é para selar com êxito o sensor de abertura. Se o sensor não é selado corretamente, água do mar pode obter para o calibre de tensão, aumentar a tração e eventualmente torná-lo inoperante. 29-sensores foram implantados ao longo de um experimento do mesocosmo no Bergeron (2005)64 e avaliou a quantidade de tempo que eles funcionavam. Os sensores mais antigo funcionaram há mais de 16 d, quando o experimento foi encerrado. A menor quantidade de tempo que um sensor durou foi 0,5 d. O comprimento médio de implantação bem sucedida do sensor foi-4,9 ± 3,0 dias, que inclui os sensores que foram concluídos no início. A maioria dos sensores trabalharam sobre a implantação de dois dias no ciclismo diel experimento de hipóxia.
O tempo de vida dos sensores deve ser melhorado, embora o dispositivo atual é apropriado para muitas aplicações. O selante de aquário contém ácido acético que pode aumentar a corrosão em sensores. No futuro, o silicone RTV da eletrônica não-ácido deve ser usado. Isso também pode evitar a tração linear que foi encontrado e que teve de ser retirada em pós-processamento. Presentemente, recomenda-se construir um novo sensor para cada gape executar.
A presente limitação é que a SGM só foi usado em um ambiente de laboratório e cabos dos sensores precisam ser feitas suficientemente longo para chegar os vários aquários. Outra limitação do método é que as análises não são feitas automaticamente e tem que ser executada com a mão e a tração linear precisa ser corrigida individualmente. Se o espaço real gape os bivalves é para ser medido, cada sensor tem que ser calibrado individualmente, como a magnitude do sinal varia de sensor para sensor. É importante no final de uma corrida para acionar os bivalves para perto de obter a magnitude e direção de spike o fechamento. Se o gape é medido no campo, condições ambientais adversas, tais como períodos de condições de baixo oxigênio podem ser detectadas pelo monitoramento os bivalves usando o SGM e detectando quando mais de um terço fechar simultaneamente.
Antes de Porter e Breitburg (2016)63, gape válvula não tinha sido medido em moluscos bivalves expostas à hipoxia diel-ciclismo. No entanto, várias técnicas existem para medir a abertura da válvula. Uma vantagem desta técnica é que é muito barato e não exige a linha direta de visão para os bivalves. Custos recorrentes são apenas os custos para extensómetros. O sistema também pode ser feito destacáveis em campo pela carcaça da ponte de Wheatstone e o datalogger em um compartimento à prova d'água para ser removido periodicamente para baixar dados. Os bivalves iria ser alojados num compartimento molhado.
Demonstramos a válvula significativa gape respostas de ostras hipóxia diel-ciclismo e demonstrar que as ostras não respondem a mudanças cíclicas no pH. Futuros trabalhos devem focar melhor selagem o sensor para reduzir falha do sensor de início ainda mais e deriva possivelmente usando a fita adesiva de calor flexível-derrete como em Jou et al (2013) 57 e usando silicone de RTV de grau eletrônico não-ácido. Estudos futuros devem medir a resposta do gape válvula de outras espécies de bivalves à hipoxia diel-ciclismo e também analisar o efeito de diel-ciclismo hipóxia e comida de limitação sobre o comportamento de abertura da válvula. É conhecido que bivalves fecham em resposta a concentrações de pouca comida, mas a interação entre hipóxia diel-ciclismo e concentração de comida não foi examinada.
Divulgações
Não temos nada para divulgar.
Agradecimentos
Agradecemos a Melinda Forseth para tirar fotos de ostras e medir sua largura de abertura no ImageJ. Agradecemos Denise Breitburg acesso para os aquários com diel ciclismo hipóxia e as condições de pH cíclico. Agradecemos o centro de investigação ambiental Smithsonian, Edgewater, Maryland, espaço para os experimentos. Os experimentos de hipóxia foram financiados por uma Administração Nacional Oceânica e atmosférica - centro para bolsa de investigação de oceano costeiro patrocinado não. NA10NOS4780138 e o fundo de Hunterdon Smithsonian para Denise Breitburg. A válvula gape medições durante a hipóxia experimentos foram financiados por uma concessão de realce do corpo docente pela Universidade de Washington para Elka T. Porter.
Materiais
| Name | Company | Catalog Number | Comments |
| Campbell CR 10x data logger | Campbell Scientific, Logan, Utah | Or other data logger. At Campbell the CR 10X has been replaced with the CR 1000 | |
| Campbell CR 10x multiplexer | Campbell Scientific, Logan, Utah | Data logger needs to have space for 12 channels | |
| Dsub connector male crimp pins | TE Connectivity | 205089-1 | pins for gape sensor leads |
| PCA tape | Micro Measurements Corp, NC | To seal the strain gauge | |
| Duro Quick Gel | Ace Hardware | Superglue | |
| SG13/1000-LY43 or LY41 | Omega Engineering Inc., Stanford, CT | Strain gauges | |
| 32 AWG (7/40) teflon Alpha wires | AlphaWire, Elizabeth, NJ | 2840/7 | Sensor cables, different colors are available |
| 1/16" heat shrink tubing | Qualtek | B01A3QKKO6 | To seal the leads of the sensor cable |
| Weller WES51 Analog Soldering Station | Amazon | Lots of soldering, need a good soldering iron. https://www.amazon.com/Weller-WES51-Analog-Soldering-Station/dp/B000BRC2XU/ref=sr_1_23?s=hi&ie=UTF8&qid=1505654295 &sr=1-23&keywords=soldering+iron | |
| Rosin Soldering Flux Paste | Amazon | Needed for soldering | |
| 60-40 Tin Lead Rosin Core Solder Wire | Amazon | Needed for soldering | |
| Aquarium sealant | Home Depot | Attach sensors to bivalve | |
| PC Laptop | Any old PC to run Campbell gape program | ||
| heat gun | Amazon | shrink shrink tubing | |
| Drill | Hardware store, Amazon | for twisting wires to make sensor cables | |
| AC to DC power module | Acopian | DB15-30 | Wheatstone bridge power supply |
| Poteniometer | Clarostat | 733A | Wheatsone bridge nulling |
| isolating BNC connector | Sterren Electronics | "200-148 | Wheatstone bridge output for multimeter |
| Fused AC receptical panel module | Adam technologies | IEC-GS-1-200 | Wheatstone bridge power supply connector |
| 976 ohm 1% resistor | Vishay Dale | CMF50976R00FHEB | Wheatstone bridge resistor |
| 1 kohm 1% resistor | Vishay Dale | CMF501K0000FHEB | Wheatstone bridge resistor |
| Potentiometer scale dial | Kilo International | 462 | 10 turn dial for nulling potentiometer |
| DB25 male panel connector | TE connectivity | 1757819-8 | Data logger connector on Wheatstone bridge |
| DB25 female panel connector | TE connectivity | 1757819-8 | Sensor connector to Wheatstone bridge |
| perforated circuit board | Vector electronics | 64P44WE | circuit board for mounting of bridge components |
| enclosure | Hammond Manufacturing | 1444-29 | Enclosure for sensor readout electronics |
Referências
- Vaquer-Sunyer, R., Duarte, C. M. Thresholds of hypoxia for marine biodiversity. Proceedings of the National Academy of Sciences of the United States of America. 105 (40), 15452-15457 (2008).
- Diaz, R. J., Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science. 321, 926-929 (2008).
- Levin, L. A., Breitburg, D. L. Linking coasts and seas to address ocean deoxygenation. Nature Climate Change. 5, 401-403 (2015).
- Diaz, R. J., Rosenberg, R. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanography and Marine Biology: An annual Review. 33, 245-303 (1995).
- Patterson, H. K., Boettcher, A., Carmichael, R. H. Biomarkers of dissolved oxygen stress in oysters: a tool for restoration and management efforts. PLoS One. 9 (8), 104440(2014).
- Altieri, A. H., Gedan, K. B. Climate change and dead zones. Global Change Biology. 21 (4), 1395-1406 (2015).
- Tyler, R. M., Brady, D. C., Targett, T. E. Temporal and spatial dynamics of diel-cycling hypoxia in estuarine tributaries. Estuaries and Coasts. 32 (1), 123-145 (2009).
- Breitburg, D. L., et al. Landscape-level variation in disease susceptibility related to shallow-water hypoxia. PLoS One. 10 (2), 0116223(2015).
- Stickle, W. B., Kapper, M. A., Liu, L. -L., Gnaiger, E., Wang, S. Y. Metabolic adaptations of several species of crustaceans and molluscs to hypoxia: toterance and microcalometric studies. Biological Bulletin. 177 (2), 303-312 (1989).
- Gamenick, I., Jahn, A., Vopel, K., Guiere, O. Hypoxia and sulphide as structuring factors in a macrozoobenthic community on the Baltic Sea shore: colonization studies and tolerance experiments. Marine Ecology Progress Series. , 73-85 (1996).
- Modig, H., Olafsson, E. Responses of Baltic benthic invertebrates to hypoxic events. Journal of Experimental Marine Biology and Ecology. 229 (1), 133-148 (1998).
- Riedel, B., Zuschin, M., Stachowitsch, M. Tolerance of benthic macrofauna to hypoxia and anoxia in shallow coastal seas: a realistic scenario. Marine Ecology Progress Series. 458, 39-52 (2012).
- Lombardi, S. A., Harlan, N. P., Paynter, K. T. Survival, acid-base balance, and gaping responses of the Asian Oyster C. ariakensis and the Eastern Oyster C. virginica during clamped emersion and hypoxic immersion. Journal of Shellfish Research. 32 (2), 409-415 (2013).
- Jansson, A., Norkko, J., Dupont, S., Norkko, A. Growth and survival in a changing environment: Combined effects of moderate hypoxia and low pH on juvenile bivalve Macoma balthica. Journal of Sea Research. 102, 41-47 (2015).
- Gooday, A. J., et al. Faunal responses to oxygen gradients on the Pakistan margin: A comparison of foraminiferans, macrofauna and megafauna. Deep Sea Research Part II: Topical Studies in Oceanography. 56 (6-7), 488-502 (2009).
- Montagna, P. A., Ritter, C. Direct and indirect effects of hypoxia on benthos in Corpus Christi Bay, Texas, U.S.A. Journal of Experimental Marine Biology and Ecology. 330 (1), 119-131 (2006).
- Villnas, A., Norkko, J., Lukkari, K., Hewitt, J., Norkko, A. Consequences of increasing hypoxic disturbance on benthic communities and ecosystem functioning. PLoS One. 7 (10), 44920(2012).
- Breitburg, D. Effects of hypoxia, and the balance between hypoxia and enrichment on coastal fishes and fisheries. Estuaries. 25 (4), 767-781 (2002).
- Costantini, M., et al. Effect of hypoxia on habitat quality of striped bass (Morone saxatilis) in Chesapeake Bay. Canadian Journal of Fisheries and Aquatic Sciences. 65 (5), 989-1002 (2008).
- Ludsin, S. A., et al. Hypoxia-avoidance by planktivorous fish in Chesapeake Bay: Implications for food web interactions and fish recruitment. Journal of Experimental Marine Biology and Ecology. 381, 121-131 (2009).
- Zhang, H., et al. Hypoxia-driven changes in the behavior and spatial distribution of pelagic fish and mesozooplankton in the northern Gulf of Mexico. Journal of Experimental Marine Biology and Ecology. 381, 80-91 (2009).
- Sparks, B. L., Strayer, D. L. Effects of low dissolved oxygen on juvenile Elliptio complanata (Bivalvia:Unionidae). Journal of the Norther American Benthological Society. 17, 129-134 (1998).
- Llanso, R. J. Effects of hypoxia on estuarine benthos: the lower Rappahannock River (Chesapeake Bay), a case study. Estuarine, Coastal and Shelf Science. 35 (5), 491-515 (1992).
- Riedel, B., Zuschin, M., Haselmair, A., Stachowitsch, M. Oxygen depletion under glass: Behavioural responses of benthic macrofauna to induced anoxia in the Northern Adriatic. Journal of Experimental Marine Biology and Ecology. 367 (1), 17-27 (2008).
- Riedel, B., et al. Effect of hypoxia and anoxia on invertebrate behaviour: Ecological perspectives from species to community level. Biogeosciences. 11 (6), 1491-1518 (2014).
- Breitburg, D. L. Near-shore hypoxia in the Chesapeake Bay: Patterns and relationships among physical factors. Estuarine Coastal and Shelf Science. 30, 593-609 (1990).
- Baumann, H., Wallace, R. B., Tagliaferri, T., Gobler, C. J. Large natural pH, CO2 and O2 fluctuations in a temperate tidal salt marsh on diel, seasonal, and interannual time scales. Estuaries and Coasts. 38, 220-231 (2015).
- Breitburg, D. L., et al. Landscape-level variation in disease susceptibility related to shallow-water hypoxia. PLoS One. 10 (2), 0116223(2015).
- de Zwart, D., Kramer, J. M., Jenner, H. A. Practical experiences with the biological early warning system "mosselmonitor". Environmental Toxicology and Water Quality. 10 (4), 237-247 (1995).
- Kadar, E., et al. Avoidance responses to aluminum in the freshwater bivalve Anodonta cygnea. Aquatic Toxicology. 55, 137-148 (2001).
- Soliman, M. F. M., El-Shenawy, N. S., Tadros, M. M., Abd El-Azeez, A. A. Impaired behavior and changes in some biochemical markers of bivalve (Ruditapes decussatus) due to zinc toxicity. Toxicological & Environmental Chemistry. 97 (5), 674-686 (2015).
- Shumway, S. E., Cucci, T. L. The effects of the toxic dinoflagellate Protogonyaulax tamarensis on the feeding and behaviour of bivalve molluscs. Aquatic Toxicology. 10, 9-27 (1987).
- Basti, L., et al. Effects of the toxic dinoflagellate Heterocapsa circularisquama on the valve movement behaviour of the Manila clam Ruditapes philippinarum. Aquaculture. 291 (1-2), 41-47 (2009).
- Tran, D., Haberkorn, H., Soudant, P., Ciret, P., Massabuau, J. -C. Behavioral responses of Crassostrea gigas exposed to the harmful algae Alexandrium minutum. Aquaculture. 298 (3-4), 338-345 (2010).
- Shumway, S. E., Koehn, R. K. Oxygen consumption in the American oyster Crassostrea virginica. Marine Ecology Progress Series. 9, 59-68 (1982).
- Nicastro, K. R., Zardi, G. I., McQuaid, C. D., Pearson, G. A., Serrao, E. A. Love thy neighbour: group properties of gaping behaviour in mussel aggregations. PLoS One. 7 (10), (2012).
- Dowd, W. W., Somero, G. N. Behavior and survival of Mytilus. congeners following episodes of elevated body temperature in air and seawater. Journal of Experimental Biology. 216 (3), 502-514 (2013).
- Higgins, P. J. Effects of food availability on the valve movements and feeding behavior of juvenile Crassostrea virginica (Gmelin). I. Valve movements and periodic activity. Journal of Experimental and Experimental Marine Biology and Ecology. 45, 229-244 (1980).
- Riisgård, H. U., Lassen, J., Kittner, C. Valve-gape response times in mussels (Mytilus edulis)-Effects of laboratory preceding-feeding conditions and in situ tidally induced variation in phytoplankton biomass. Journal of Shellfish Research. 25, 901-911 (2006).
- Robson, A. A., De Leaniz, C. G., Wilson, R. P., Halsey, L. G. Behavioural adaptations of mussels to varying levels of food availability and predation risk. Journal of Molluscan Studies. 76, 348-353 (2010).
- Robson, A. A., de Leaniz, C. G., Wilson, R. P., Halsey, L. G. Effect of anthropogenic feeding regimes on activity rhythms of laboratory mussels exposed to natural light. Hydrobiologia. 655, 197-204 (2010).
- Nicastro, K. R., Zardi, G. I., McQuaid, C. D., Stephens, L., Radloff, S., Blatch, G. L. The role of gaping behaviour in habitat partitioning between coexisting intertidal mussels. BMC Ecology. 10, 17(2010).
- Loosanoff, V. S., Nomejko, C. A. Feeding of oysters in relation to tidal stages and to periods of light and darkness. Biological Bulletin. 90 (3), 244-264 (1946).
- Comeau, L. A., Mayrand, E., Mallet, A. Winter quiescence and spring awakening of the Eastern oyster Crassostrea virginica at its northernmost distribution limit. Marine Biology. 159 (10), 2269-2279 (2012).
- Pynonnen, K. S., Huebner, J. Effects of episodic low pH exposure on the valve movements of the freshwater bivalve Anodonta cygnea L. Water Research. 29 (11), 2579-2582 (1995).
- Jakubowska, M., Normant-Saremba, M. The effect of CO2-induced seawater acidification on the behaviour and metabolic rate of the baltic clam Macoma balthica. Annales Zoologici Fennici. 52 (5-6), 353-367 (2015).
- Jakubowska, M., Normant, M. Metabolic rate and activity of blue mussel Mytilus edulis trossulus.under short-term exposure to carbon dioxide-induced water acidification and oxygen deficiency. Marine and Freshwater Behaviour and Physiology. 48 (1), 25-39 (2015).
- Newell, C. R., Wildish, D. J., MacDonald, B. A. The effects of velocity and seston concentration on the exhalent siphon area, valve gape and filtration rate of the mussel Mytilus edulis. Journal of Experimental Marine Biology and Ecology. 262, 91-111 (2001).
- Maire, O., Amouroux, J. -M., Duchene, J. -C., Gremare, A. Relationship between filtration activity and food availability in the Mediterranean mussel Mytilus galloprovincialis. Marine Biology. 152 (6), 1293-1307 (2007).
- Borcherding, J. Ten years of practical experience with the Dreissena-monitor, a biological early warning system for continuous water quality monitoring. Hydrobiologia. 556 (1), 417-426 (2006).
- Frank, D. M., Hamilton, J. F., Ward, E. E., Shumway, S. E. A fiber optic sensor for high resolution measurement and continuous monitoring of valve gape in bivalve molluscs. Journal of Shellfish Research. 26 (2), 575-580 (2007).
- Wilson, R., Reuter, P., Wahl, M. Muscling in on mussels: New insights into bivalve behaviour using vertebrate remote-sensing technology. Marine Biology. 147 (5), 1165-1172 (2005).
- Nagai, K., Honjo, T., Go, J., Yamashita, H., Seok Jin, O. Detecting the shellfish killer Heterocapsa circularisquama (Dinophyceae) by measuring bivalve valve activity with a Hall element sensor. Aquaculture. 255 (1-4), 395-401 (2006).
- Robson, A., Wilson, R., de Leaniz, C. G. Mussels flexing their muscles: a new method for quantifying bivalve behaviour. Marine Biology. 151 (3), 1195-1204 (2007).
- Robson, A. A., Thomas, G. R., de Leaniz, C. G., Wilson, R. P. Valve gape and exhalant pumping in bivalves: optimization of measurement. Aquatic Biology. 6 (1-3), 191-200 (2009).
- de Zwart, D., Kramer, J. M., Jenner, H. A. Practical experiences with the biological early warning system "mosselmonitor". Environmental Toxicology and Water Quality. 10 (4), 237-247 (1995).
- Jou, L. -J., Lin, S. -C., Chen, B. -C., Chen, W. -Y., Liao, C. -M. Synthesis and measurement of valve activities by an improved online clam-based behavioral monitoring system. Computers and Electronics in Agriculture. 90, 106-118 (2013).
- Barile, N. B., Scopa, M., Recchi, S., Nerone, E. Biomonitoring of coastal marine waters subject to anthropogenic use: development and application of the biosensor Mosselmonitor. Ovidius University Annals of Chemistry. 27 (2), 81-86 (2016).
- Ballesta-Artero, I., Witbaard, R., Carroll, M. L., van der Meer, J. Environmental factors regulating gaping activity of the bivalve Arctica islandica in Northern Norway. Marine Biology. 164 (5), 116(2017).
- Ekelof, S. The genesis of the Wheatstone bridge. Engineering Science and Education Journal. 10, 37-40 (2001).
- Keppel, A. G., Breitburg, D. L., Wikfors, G. H., Burrell, R. B., Clark, V. M. Effects of co-varying diel-cycling hypoxia and pH on disease susceptibility in the eastern oyster Crassostrea virginica. Marine Ecology Progress Series. 538, 169-183 (2015).
- Burrell, R. B., Keppel, A. G., Clark, V. M., Breitburg, D. L. An automated monitoring and control system for flow-through co-cycling hypoxia and pH experiments. Limnology and Oceanography: Methods. 14, 168-185 (2015).
- Porter, E. T., Breitburg, D. L. Eastern oyster Crassostrea virginica, valve gape behavior under diel-cycling hypoxia. Marine Biology. 163 (218), (2016).
- Bergeron, C. M. The impact of sediment resuspension on mercury cycling and the bioaccumulation of methylmercury into benthic and pelagic organisms. , University of Maryland. M.S. thesis (2005).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados