
Method Article
Un modèle de blessure de Drosophila In Vivo pour étudier la neurogenèse dans le périphérique et le système nerveux Central
Dans cet article
Résumé
Nous présentons ici un protocole utilisant la drosophile neurone sensitif - modèle de blessure de neurone arborisation dendritique (da), qui combine en vivo live imaging, deux photons laser axotomie/dendriotomy et la puissante mouche génétique boîte à outils, comme une plateforme de criblage éventuels promoteurs et inhibiteurs de la neurogenèse.
Résumé
La capacité de régénération des neurones endommagés régit la neurogenèse et la récupération fonctionnelle après un traumatisme du système nerveux. Dans les dernières décennies, diverses intrinsèques et extrinsèques inhibiteurs facteurs impliqués dans la restriction de la régénération axonale ont été identifiés. Toutefois, simplement supprimer ces signaux inhibiteurs est insuffisant pour une saine régénération, indiquant l’existence de mécanismes réglementaires supplémentaires. Drosophila melanogaster, la mouche à fruit, partage des gènes évolutivement conservés et voies de signalisation avec les vertébrés, y compris les humains. Alliant la puissante boîte à outils génétique des mouches à deux photons laser axotomie/dendriotomy, nous décrivons ici le neurone sensitif Drosophila – modèle de blessure neurone arborisation dendritique (da) comme une plate-forme pour le dépistage systématique pour roman régulateurs de régénération. Brièvement, ce paradigme inclut un) la préparation de larves, l’induction b) lésion à dendrite(s) ou en utilisant un laser deux photons, c) direct confocal imagerie après lésion d) analyse de données et d’axon(s). Notre modèle permet une blessure très reproductible de neurones marqués, les axones et les dendrites des sous-types neuronales bien définis, dans les périphériques et le système nerveux central.
Introduction
L’incapacité des axones à se régénérer après une lésion du système nerveux central (CNS), peut conduire à une invalidité permanente chez les patients et joue également un rôle dans les déficits neurologiques irréversibles dans des maladies neurodégénératives1,2 ,3,4,5. L’environnement de la CNS, ainsi que la capacité de croissance intrinsèque des neurones, détermine si les axones sont capables de régénérer après un traumatisme. Les facteurs extracellulaires des oligodendrocytes, astroglials et fibroblastiques sources auraient dû être divulgués d’entraver la croissance neuronale4,6,7,8, mais l’élimination de ces molécules ne permet que de limited5la germination. Intrinsèque de la régénération de signaux peuvent influencer le succès régénératrice5,9 et représentent des cibles thérapeutiques potentielles, mais ces processus ne sont toujours pas bien définis au niveau moléculaire. Augmentations de facteur trophique de signalisation ou d’élimination des freins endogènes, tels que le gène Pten phosphatase10, peuvent entraîner la régénération axonale dans certaines circonstances. Combinaisons de différentes méthodes individuellement efficaces fournissent également seulement restauration globale limitée à ce jour11,12,13,14. Par conséquent, il y a un besoin désespéré d’identifier des voies supplémentaires pour thérapie ciblée. En plus de l’ouverture de la repousse axonale, si et comment les axones recâbler à la bonne cible, la spécificité de la réforme de synapse et atteindre la récupération fonctionnelle d’importantes questions sans réponse.
En résumé, notre compréhension actuelle de la machinerie dictant la régénération axonale est encore très fragmentaire. Une partie du problème est la difficulté technique de l’étude axon régénération chez les mammifères en temps réel, une approche qui est coûteux, chronophages et stimulant pour la réalisation des écrans génétiques à grande échelle. Drosophila melanogaster, en revanche, s’est avéré pour être un système extrêmement puissant pour l’étude des questions biologiques complexes. La drosophile a joué un rôle important dans la définition des gènes et de signalisation des voies qui sont remarquablement conservées chez les humains et a été un modèle pour l’étude des conditions humaines, telles que les maladies neurodégénératives, grâce aux outils de la génétique moléculaire vaste disponibles pour manipuler les gènes fonction15. En particulier, les mouches des fruits sont considérés comme un outil idéal pour la découverte de gènes impliqués dans la lésion neurale et repousse le15,16. Plusieurs modèles de mouche lésion neuronale ont été développés, y compris la tête adulte ou larvaire nerveuse ventrale cordon (VNC) poignardant avec aiguilles, VNC larvaire ou écrasement du nerf avec forceps, neurone larvaire laser axotomie, enlèvement des neurones récepteurs olfactifs, explants de traumatisme crânien, et lésion des nerfs périphériques par aile départ15,17,18,19,20,21,22,23. Passionnante, récent travail utilisant modèles de lésion de drosophile ont fait progresser notre compréhension des voies cellulaires et génétiques utilisées par le système nerveux pour répondre aux blessures neurales, dont certains auraient dû être divulgués à conserver chez les mammifères24 ,,25. Encore une fois, cela fait ressortir l’utilité de cet organisme modèle pour identifier de nouveaux mécanismes de réparation neuronale.
Décrit ici est un modèle de blessure du Drosophila biphotonique par laser larvaire neurone sensitif. Un laser à deux photons a été utilisé pour couper des axones dans le poisson-zèbre en vivo en 200326. La même année, le premier dendriotomy de laser a été réalisée chez la drosophile à l’aide d’un de laser pulsé azote27. Lasers femtosecondes a été utilisé pour établir des modèles de régénération axonale28peu après, plusieurs laboratoires de c. elegans . En 2007, Wu et ses collègues comparativement et signalé les différences entre les blessures de laser chez c. elegans induite par différents types de lasers à29. En 2010, la régénération axonale après laser axotomie fut montrée chez Drosophila30. S’appuyant sur cette littérature de blessure laser vaste, nous avons développé un modèle de mouche blessures neurales à l’aide de deux photons laser, ce qui permet à induction précise des lésions aux sites ciblés avec une perturbation minimale des tissus voisins, fournissant un endroit relativement propre système pour étudier les propriétés intrinsèques et extrinsèques de neurodégénérescence avec résolution de cellule unique. Plus précisément, nous avons établi un ensemble de méthodes de blessures pour les neurones sensoriels de l’arborisation dendritique (da) dans les deux système nerveux périphérique (PNS) et CNS. Neurones dopaminergiques peuvent être regroupés en quatre classes distinctes se distingués principalement par leur complexité de ramification de dendrite : classe I à IV,31. Nos travaux publié montrent que la régénération des neurones da ressemble à modèles de lésion chez les mammifères au niveau phénotypique et moléculaire : neurones dopaminergiques afficher des propriétés de régénération spécifiques de classe, avec la classe IV, mais pas de classe I ou les neurones da III présentant la régénération dans le PNS ; axones de neurones classe IV da régénèrent robuste dans la périphérie, mais leur potentiel de régénération est considérablement réduite dans le SNC, ressemblant ainsi à des neurones ganglionnaires (DRG) racine dorsale chez les mammifères ; activité de mTOR via Pten suppression ou Akt surexpression améliore la régénération axonale dans la volée CNS19. En utilisant ce modèle de blessures, nous ont de bons écrans génétiques et ont identifié l’enzyme de traitement RNA Rtca comme un facteur inhibiteur évolutivement conservé pour la régénération axonale, reliant des lésions axonales au stress cellulaire et modification de RNA20 .
Dans le modèle présenté, la blessure est induite par laser axotomie/dendriotomy des larve classe IV ou les neurones da III, étiquetés par ppk-CD4-tdGFP ou 19-12-Gal4, SAMU-CD4-tdGFP, repo-Gal80, respectivement. La blessure s’effectue sur 2nd à 3rd stade larvaire à environ 48 à 72 heures après la ponte (h AEL). Pour PNS axotomie la lésion est visée à la section de l’axone ~ 20-50 µm loin du corps de la cellule, pour axotomie CNS d’une superficie d’environ 20 µm de diamètre à la jonction de la commissure dans VNC et dendriotomy aux points direction dendritiques primaires. Même neurone est imagé à 8 à 24 heures après la lésion (AI) pour confirmer la transection complète et à 48-72 h AI pour évaluer la régénération. Par le biais de l’imagerie confocale Time-lapse, la dégénérescence et la régénération des axones/dendrites individuels qui ont été blessés en vivo peuvent être surveillés au fil du temps.
Protocole
1. préparation des plaques de Culture et bouteilles
- Préparation des boîtes de gélose de jus de raisin
- Ajouter 10 g de poudre d’agar, jus de raisin 200 mL et mL 192 ddH2O dans un bécher et four à micro-ondes pendant environ 4-5 min, en remuant par intermittence jusqu'à dissolution complète de la gélose.
- Sous une hotte, refroidir la solution à environ 60 ° C. Ajouter 4,2 mL d’éthanol à 95 % et 4,0 mL d’acide acétique glacial. Réglez le volume total de la solution à 400 mL avec FD2O. Mix bien.
- Pour chaque plaque de 35 mm, ajouter environ 2-3 mL de la solution. Faire environ 120-150 plaques pour un total de 400 mL de solution d’agar jus de raisin.
- Attendre 10 min pour refroidir les plaques et solidifier la solution d’agar. Emballez le jus de raisin géloses dans des sacs ziplock auto scellés et conserver à 4 ° C pour une utilisation future.
- Préparation des flacons de culture de drosophile
- Utilisez une lame pour perforer un trou triangulaire 1,5 cm de longueur côté sur une paroi de la bouteille de culture de drosophile et remplissez le trou avec une boule de diamètre de 2 à 2,5 cm de coton pour la ventilation.
- Brancher la bouteille avec une gélose au jus de raisin additionnée de 0,5 cm3 de la pâte de levure.
2. collection de larves de drosophile
- Mettre en place des croisements de mouches adultes
- Placer 10 femelles vierges et 5 mâle adulte vole ensemble dans un flacon d’hémoculture branché avec une gélose au jus de raisin.
- Placer le fond de la bouteille vers le haut à 25° C, afin que les mouches pondront sur la gélose au jus de raisin. Changer la plaque avec la pâte de levure au moins une fois par jour. Pour une période d’échantillonnage de 2 h, qui permet la récolte de larves d’un stade de développement homogène, commencez avec plus de 20 femelles vierges et 10 mâles à l’étape 2.1.1.
- La plaque à 25° C dans un plat de Pétri de 60 mm avec un chiffon humide, par exemple, trempée dans une solution acide propionique 0,5 % de la culture. Organiser le tissu afin qu’il ne bloque pas l’alimentation en oxygène.
Remarque : La solution de l’acide propionique est utilisée pour maintenir l’humidité dans le plat et éviter la croissance de moisissures.
- Récolte de larves d’un âge spécifique
- Utilisez une paire de pinces pour transférer les larves de la scène désirée, par exemple, 2ème à la 3rd stade larvaire à 48-72 h après la ponte (AEL), pour une nouvelle gélose au jus de raisin sans pâte de levure.
- Supprimer la levure de peau de la larve en lui faisant ramper sur la nouvelle plaque, pour empêcher d’éventuels interférer de levure avec une axotomie laser et l’imagerie. Vous pouvez également nettoyer soigneusement les larves en les lavant dans un plat de PBS et asséchant brièvement sur un morceau de papier de soie.
3. deux photons blessures et imagerie confocale
- Configuration de microscope
Remarque : Un confocal laser scanning microscope avec un laser à deux photons a été utilisé pour cette expérience, mais les autres systèmes avec une configuration équivalente aussi suffira. Le laser à deux photons (930 nm) a été utilisé pour la livraison des blessures et un laser à l’Argon (488 nm) a été utilisée pour l’imagerie confocale de GFP.- Au début de chaque session, allumez le deux photons laser et/ou le laser(s) confocale et le microscope. Ouvrez le logiciel d’imagerie.
- Pour blessures de deux photons, définir les paramètres suivants pour l’imagerie des GFP au laser biphotonique à 930 nm (1 950 mW).
- Sélectionnez le mode de balayage de ligne. Ouvrir complètement le trou d’épingle. Augmenter l’intensité du laser à environ 20 % (390 mW).
- Sélectionnez 512 x 512 comme le balayage de trame. Utiliser la vitesse de balayage maximale (généralement avec le temps de pause de pixel à 0,77 µs). S’assurer que le nombre moyen est de 1 et bits de profondeur de 8 bits.
- Réglage gain à ~ 750 et l’offset 0.
- Enregistrez ce protocole expérimental prédéterminé sous 2p GFP 930 Ablation, permettant la réutilisation simple des expériences à l’avenir.
- Pour l’imagerie confocale, définir les paramètres suivants pour l’imagerie des GFP au laser Argon à 488 nm :
- Sélectionnez l’onglet Acquisition , puis Z-pile.
- Sous «Laser», mettre en marche pour le laser à Argon 488 nm.
- Aller vers les canaux, sélectionnez le laser de 488 mm et augmenter la puissance de laser à 5 à 10 %. Pour le trou d’épingle, utiliser l’unité aéré 1-2 (UA). Ajuster le gain à 650.
- En Mode d’Acquisition, sélectionnez 1024 x 1024 comme l’analyse de l’image, utilisez la vitesse de balayage maximale, un nombre moyen de 2 et les bits de profondeur de 8 bits.
- Enregistrez ce protocole expérimental prédéterminé sous Imagerie GFP.
- Anesthésie de larves avec l’éther diéthylique et montage
- Sous une hotte, placez un plat de verre de 60 mm en plastique 15 cm boîte de Pétri. Plier et poser un morceau de papier de soie dans le bas du verre plat, puis placez une gélose au jus de raisin sur le tissu. Ajouter l’éther dans le plat de verre, au point où le papier de soie est trempé et il y a une couche d’éther liquide restant dans le plat. Garder le couvercle à tout moment.
- Préparer une lame avec une goutte d’halocarbure 27 huile au centre. Ajouter 4 taches de graisse sous vide sur les quatre coins de la diapositive, pour soutenir plus tard la lamelle.
- Forceps permet de ramasser une larve nettoyée et placez-le sur la gélose dans le plat de verre de 60 mm. Couvrir le plat en verre avec son couvercle et attendez que la larve cesse de bouger. Pour PNS blessure/imagerie, sortez la larve dès que sa queue s’arrête à secousses. Pour le CNS, attendre que la larve entière devient immobile, en particulier les segments de tête.
Remarque : Le moment de l’exposition de l’éther est essentiel. Voir la Discussion. - Soigneusement, ramasser la larve anesthésiée et placez-le en tête-debout dans la goutte de Huilehalocarbone sur la diapositive. Ajouter un lamelle couvre-objet sur le dessus de la diapositive. Une pression douce permet d’appuyer sur la lamelle couvre-objet, jusqu'à ce qu’il touche la larve (Figure 1 a).
- Décaler de la larve en poussant doucement la lamelle vers la gauche ou la droite pour rouler la larve, afin que le neurone/axon/dendritique d’intérêt est sur le haut et le plus proche de la lentille du microscope.
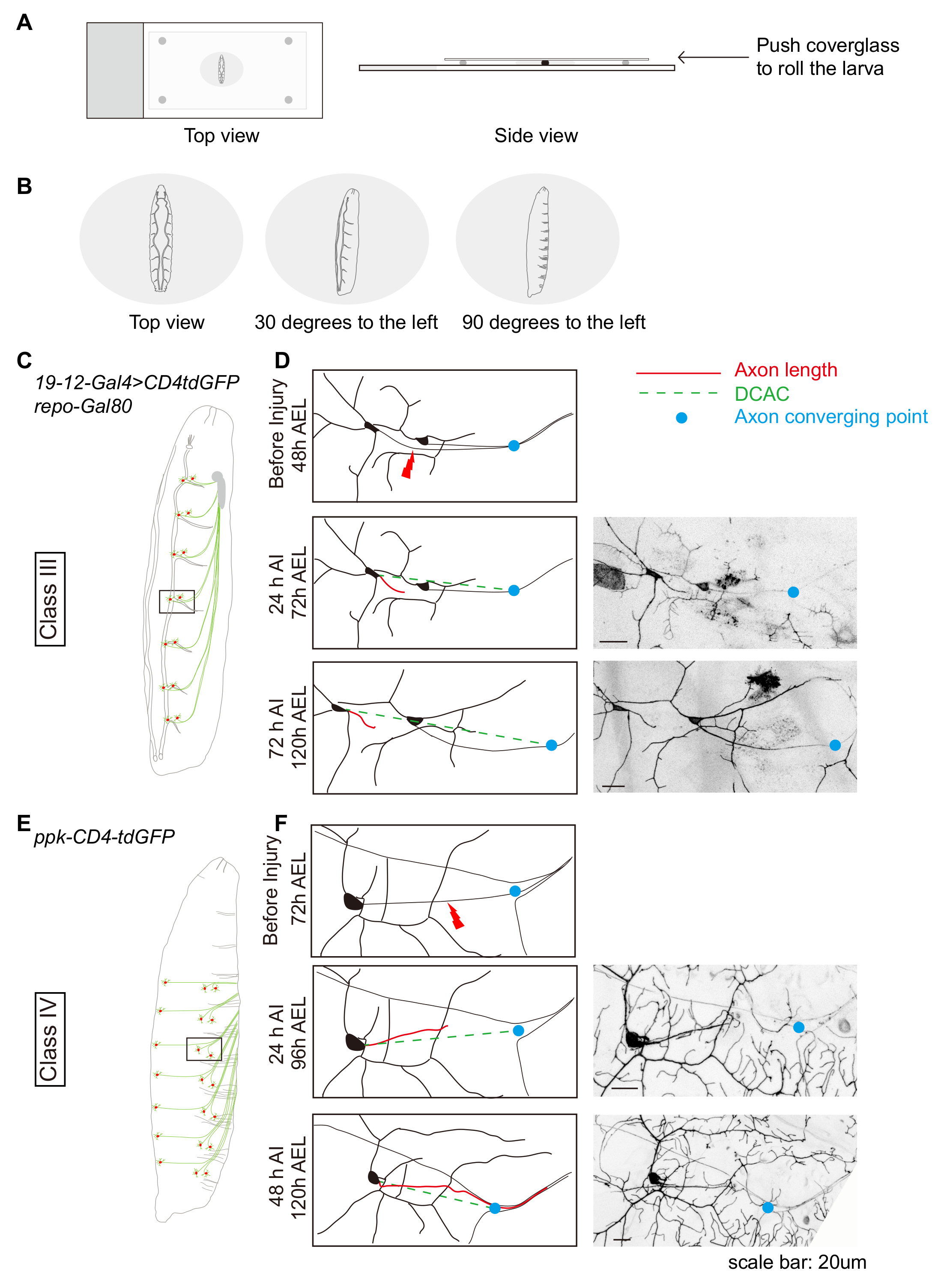
- Des blessures PNS, monter la face dorsale de la larve, afin que les deux les trachées sont visibles. Puis rouler la larve ~ 30 degrés vers la gauche pour blesser les axones de neurones classe III da (Figure 1 b et 1c), 90 degrés pour blesser les axones de neurones classe IV da (Figure 1 b et 1F) ou ~ 30 degrés pour blessant classe IV da (dendrites) neuron Figure 2 a).
- Pour les blessures de la CNS, positionner la larve pour être parfaitement ventrale vers le haut (Figure 3 a), afin que la région d’intérêt est plus proche de la lentille du microscope dans le plan z.
- Blessures par deux photons laser
- Placez le chariot avec la larve sous le microscope et fixer en place avec le support de diapositive sur la scène. Objectif d’utilisation le 10 X (NA 0,3) pour trouver la larve.
- Ajouter 1 goutte d’huile objective sur la lamelle couvre-objet, passez le 40 X (NA 1,3) objectif et ajuster le focus.
- Basculez vers le mode de numérisation et de réutiliser le protocole expérimental 2p GFP 930 Ablation. Assurez-vous que le trou d’épingle est complètement ouvert.
Remarque : La configuration doit être optimisée basés sur des systèmes individuels. - Démarrer le mode Live pour localiser la région d’intérêt (ROI) et d’affiner les réglages pour obtenir la bonne qualité d’image avec le zoom approprié.
Remarque : L’objectif de cette étape est de trouver la neurone/axon/dendrite à blesser, plutôt que de prendre la meilleure qualité d’image. Par conséquent, utiliser les paramètres minimes permettant de visualiser la zone cible, afin d’éviter la surexposition ou photoblanchiment. - Arrêter le scan Live , afin que le bouton rogner deviendront disponible. Laissez l’image fixe à servir de la feuille de route. Sélectionnez le mode de récolte et ajuster les fenêtres de balayage de se concentrer sur l’objectif d’être blessés.
- Réduire le retour sur investissement pour être de la taille du foyer éventuel de la lésion. Par exemple, juste couvrir la largeur d’un axone ou d’une dendrite, pour assurer la précision de la lésion et de réduire les dommages causés aux tissus voisins. Si vous le souhaitez, effectuer un zoom avant sur le retour sur investissement avant recadrage, permettant une blessure plus précis.
- Ouvrez une nouvelle fenêtre d’imagerie. Réduire la vitesse de balayage et augmenter l’intensité du laser. Déterminer l’augmentation d’intensité laser fondée sur le signal de fluorescence de tissus scanné en mode Live .
- En général, définissez l’intensité du laser biphotonique à partir de 25 % pour les blessures de la PNS et 50 à 100 % pour les blessures VNC. Pour blessures PNS d’axon, assurez-vous que l’intensité du laser est ~ 480 mW et pixel temporisation est 8,19 μs. Pour la blessure d’axon VNC, veiller à ce que le temps de pause laser intensité et pixel sont habituellement 965-1930 mW et 8.19-32,77 μs, respectivement.
- Commencer le balayage continu . Laisser le curseur plane sur le bouton continue . Garder un œil sur l’image et arrêter le scan dès qu’on observe une augmentation drastique de fluorescence.
Remarque : L’apparition de l’épi de la fluorescence est due à auto-fluorescence au site de lésion. - Basculez vers le mode direct en réutilisant les paramètres. Trouver la région d’intérêt qui visait simplement en ajustant la mise au point.
Remarque : Une bonne indication de blessure réussie est l’apparition d’un petit cratère, anneau-comme la structure ou des débris localisés au site de lésion. - Déplacer vers le neurone suivant et répétez l’étape 3.3.5, de blesser plusieurs neurones chez un seul animal. Ou répéter l’étape 3.3.5 tandis que graduellement augmenter la puissance et/ou la réduction de vitesse de balayage si la lésion initiale était insuffisante.
NOTE : Dans le cas où la puissance du laser est trop élevée, une grande surface endommagée sera visible dans l’image scan direct après l’accident. Trop dommage peut causer la mort de la larve. - Récupérer la larve en soigneusement retirer la lamelle et en transférant la larve blessée sur une plaque de nouveau avec la pâte de levure. Plusieurs grottes de fossé sur la gélose avec une pince ; Sinon, faire une île d’agar dans la plaque au lieu d’utiliser la plaque entière, afin de réduire la possibilité de la larve ramper hors de la plaque.
- Mettre la plaque dans un plat de Pétri de 60 mm avec le tissu humide (imbibé de solution d’acide propionique de 0,5 %) et de la culture à température ambiante ou 25° C.
Remarque : La larve reste au stade larvaire pendant environ une journée supplémentaire à température ambiante (22° C) par rapport à 25° c.
- Imagerie confocale après l’accident
- Image de la larve blessée aux moments souhaités par la préparation de la larve en utilisant la même procédure d’anesthésie et de montage comme dans l’étape 3.2, puis d’imagerie au laser confocale.
Remarque : L’Image la larve après 24 h après la lésion (AI) pour confirmer les lésions axonales et à 48 h AI (neurones dopaminergiques de classe IV) ou 72 h AI (neurones dopaminergiques de classe III) pour évaluer la régénération. - Localiser la larve à l’aide de l’objectif 10 X, puis passez à un 25 X (NA 0,8) objectif. Réutiliser le protocole expérimental GFP Imaging.
- Cliquez sur le bouton Live et trouver le même neurone blessé auparavant.
- Définir la première et la dernière position de Z dans la fenêtre de scan direct. Appuyez sur stop, puis cliquez sur Start expérimenter pour acquérir une image de Z-pile.
Remarque : Assurez-vous qu’un point de la normalisation (le point de convergence de l’axone) est inclus lors de la capture d’images afin que la quantification de la régénération est possible (Figure 1, 1F) – Ceci est discuté plus en détail dans la section analyse de données. - Passer au Traitement d’images, sélectionnez l’image vient d’être prise et générer une projection de l’intensité maximale. Économiser le z-pile et intensité maximale des images de projection.
- Image de la larve blessée aux moments souhaités par la préparation de la larve en utilisant la même procédure d’anesthésie et de montage comme dans l’étape 3.2, puis d’imagerie au laser confocale.
4. analyse des données
- Traiter et quantifier des images en utilisant le logiciel d’imagerie ou ImageJ.
- Quantification de la régénération axonale dans le PNS
- Calculer le pourcentage de régénération, qui se réfère au pourcentage de régénération des axones parmi tous les axones qui ont été lésés. Marquer un axone comme régénérant tant qu’il repousse au-delà du site de la lésion.
- Mesurer la longueur de la régénération, qui est l’augmentation de la longueur de l’axone. Si quantifier avec ImageJ, utilisez l’outil de Courbe segmentée pour retracer l’axone régénérée et utiliser la mesure dans le menu déroulant analyser pour obtenir la longueur de la ligne représentant l’axone régénérée.
- Calculer l' Indice de régénération, qui est l’augmentation de longueur normalisée axon.
Remarque : Longueur de l’axone est normalisé par la distance entre le corps cellulaire et l’axone convergeant point (DCAC) – axon longueur/DCAC (Figure 1 et 1F). Cette valeur permet de compte pour toute la croissance axonale c’est en raison de la mise à l’échelle larvaire. Une valeur positive représente la régénération, la valeur 0 ne signifie aucune régénération et une valeur négative signifie rétraction.
- Quantification de la régénération de la dendrite
- Calculer le pourcentage régénération, c'est-à-dire le pourcentage de neurones dopaminergiques parmi tous ceux rompu qui montrent repousse évidente dendrite.
NOTE : Dendrite repousse est marquée comme positifs si nouvelles dendrites repoussent hors de la tige rentrée dendritique et au-delà du site de la lésion. Le site de la lésion est déterminé par les points de repère et dans certains cas, il est facilement visible en raison de l’autofluorescence induite par la lésion résiduelle. - Calculer l' augmentation des points de ramification, qui compte l’ajout de nouveaux points de branche dendritiques après une blessure.
- Calculer l' augmentation de la longueur totale de dendrite, qui est la durée cumulative de toutes les nouvelles dendrites sont complétés par des blessures.
- Calculer le pourcentage régénération, c'est-à-dire le pourcentage de neurones dopaminergiques parmi tous ceux rompu qui montrent repousse évidente dendrite.
- Quantification de la régénération axonale dans le SNC
Remarque : Si un site de lésion montre la dégénérescence au premier temps point imagé (Figure 3 b), il figurera dans l’analyse de la durée de régénération et de taux.- Mesurer la longueur de la régénération, qui correspond à la longueur de l’axone bout.
Remarque : L’axone bout d’un site de lésion unique est identifié comme il prend sa source au large de l’itinéraire initial d’axon avant l’accident. - Calculer la longueur de régénération après normalisation, qui normalise la longueur de la régénération à la longueur du segment de la commissure - la distance longitudinale entre les commissures (« Y » dans les figures 3 a et 3 b).
Remarque : Cette valeur permet de corriger l’effet des différences de taille des larves. - Calculer le taux de régénération, c'est-à-dire le pourcentage de régénération des segments parmi tous les segments qui ont été lésés, pour refléter la capacité de régénération d’un génotype particulier.
- Mesurer la longueur de la régénération, qui correspond à la longueur de l’axone bout.
Résultats
Neurones dopaminergiques montrent régénération différentielle potentiels entre le périphérique et du système nerveux central, ainsi que la spécificité de la classe. Ceci fournit une occasion unique de dépister les nouveaux facteurs qui sont requises pour la régénération axonale (à l’aide de la classe des blessures IV PNS), ainsi que celles qui sont inhibitrices de la régénération (avec la blessure de classe IV CNS et classe blessure III PNS).
Régénération axonale dans le PNS
À titre d’exemple, la caractérisation de la régénération des neurones dopaminergiques de classe III et classe IV est décrite. Ces neurones sont situés bilatéralement dans chaque segment du corps. Plusieurs neurones peuvent être blessés dans la même larve ; en général, 3-4 neurones dans la partie droite de segments abdominaux A7-A2. Classe III et classe IV da neurones peuvent être visualisées par le 19-12-Gal4, SAMU-CD4tdGFP, repo-Gal80 et ppk-CD4tdGFP, respectivement. Une anesthésie et monter les larves AEL de 48-72 h, tel que décrit, en ajustant la position des larves afin que les neurones dopaminergiques d’intérêt soient vers le haut (Figure 1 a et 1 b). Habituellement, nous blesser la ddaF de classe III et classe IV v'ada neurones (Figure 1 et 1E). Effectuer une axotomie et récupérer des larves comme décrit. Peu de temps après la lésion (AI), les larves seront est remis de locomotion normale de chirurgie et de la pièce. Le taux de survie ici est en général plus de 80 %. Jeter les larves qui sont morts ou malades. Remonter le reste comme décrit et évaluer la dégénérescence. Après 24 h AI, les axones distales devraient avoir terminé la dégénérescence à ce temps point19, et la tige de l’axone sera visible (Figure 1 et 1F). Reimage la même larve à 48 h AI des neurones dopaminergiques de classe IV ou 72 h AI des neurones dopaminergiques de classe III pour évaluer la régénération. Habituellement, nous visons à évaluer au moins 20 neurones lésés par les conditions expérimentales. En (type sauvage) WT animaux, alors que généralement environ 70 % des da coupé classe IV les neurones seront sont régénérées au-delà du site de la lésion (Figure 1F), neurones dopaminergiques de classe III ne parviennent pas à repousser, attestent par cône de croissance au point mort (Figure 1).
Régénération de dendrite
Nous effectuons généralement dendriotomy sur la classe IV da neurones CDDA (Figure 2 a). Comme illustré dans le diagramme schématique, la blessure vise le point principale branche dendritiques. Après expérience, lorsque blessé 48 h AEL, environ 50 % des neurones CDDA régénérer leurs dendrites (Figure 2 b). Dans les 50 % restants, les dendrites voisins envahissent et couvrent l’espace vacant. En outre, le potentiel de régénération de ces neurones est réduit si blessé à un stade de développement plus tard.
Régénération axonale dans le SNC
Pour lésion axonale VNC, le taux de survie des larves varie beaucoup et dépend de l’âge au cours de laquelle les blessures sont induite. Basé sur l’expérience, des larves de 48-72 h AEL ont généralement le taux de survie plus élevé (> 60 %) parmi les différentes étapes testées. Âgés de moins de 48 h AEL larves survivent mal après une blessure, tandis que chez les plus de 72 h AEL âgés, il est difficile d’introduire des blessures dans le VNC. En outre, il est plus facile à provoquer des blessures sur les segments de la commissure postérieure de l’extrémité antérieure, comme ces segments postérieurs de la VNC sont plus proches de la surface ventrale et donc plus accessibles par laser (Figure 3 a).
Pour blesser classe IV da neurone les axones dans le SNC, monter les larves comme décrits précédemment (Figure 3 a). Sous le microscope, localiser la structure de l’échelle des faisceaux d’axones qui font partie de la VNC (Figure 3 a) et effectuer une axotomie comme indiqué. Dégénérescence est confirmée à 8 h, Amnesty International et la régénération sont évaluées à 24 et 72 h AI. Comme illustré à la Figure 3 b, axones à 8 h AI ont déjà commencé à dégénérer et à 24 h AI, la régénération axonale est observée, tandis que les débris de l’axone peut encore être trouvé autour des sites de lésions. Les axones WT afficher repousse limitée à VNC et ne parviennent pas à se reconnecter les lacunes générés par la lésion (Figure 3 b). Afin de quantifier la capacité de régénération des axones lésés, la durée de repousse après que blessure est mesurée et la longueur du segment commissure (Y dans la Figure 3 a) est utilisé pour la normalisation (Figure 3).

Figure 1 : la régénération axonale neurone de Da dans la périphérie affiche la spécificité de la classe. (A et B) Schéma montrant la position des larves. (C) schéma de neurones dopaminergiques de classe III. (D) les axones de la classe III da neurones ddaF, étiqueté avec le 19-12-Gal4, SAMU-CD4-tdGFP, repo-Gal80 / +, ne parviennent pas à repousser. (E) schéma de neurones dopaminergiques classe IV. Axones (F) de la classe IV da neurones v'ada, étiqueté avec ppk-CD4-tdGFP / +, repousse au-delà du site de lésion. (D et F) Ligne rouge indique la longueur de l’axone tandis que la ligne pointillée verte marque la distance entre le corps cellulaire et l’axone convergeant point (DCAC). Le point bleu marque le point de convergence d’axon. Echelle = 20 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 : régénération de dendrite Da neurone. (A) représentation Illustrative des neurones dopaminergiques de classe IV. (B) représentation Illustrative de la régénération de la dendrite dans la classe IV da neurone CDDA, labellisé par ppk-CD4-tdGFP / +. Ablation laser vise le point de la branche principale et est effectuée 48 h AEL. Lésion 24h AI transection de la neurite est confirmée, et 72 h AI régénération est quantifiée. Dendrites des neurones CDDA démontrent repousse importante, avec nouvelles ramifications dendritiques jaillissant de la tige sectionnée pour revêtir l’espace vacant. Il est à noter que nouvelles branches terminales sont ajoutés en permanence pour les dendrites blessés à ce stade du développement. Echelle = 20 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 3 : la régénération axonale neurone de Da dans le VNC. (A) dessin schématique d’une larve de drosophile montés sur des lamelles et imagés sous le microscope. Classe IV da neurone axones dans le VNC visualisées dans un ppk-CD4tdGFP / + larve. Deux segments de commissure candidats sont indiqués dans l’image agrandie et le dessin schématique. Chacun d’eux possède deux sites de blessures (cercles rouges). (B) les images confocales un segment lésé imagés à 8, 24 et 72 h après la lésion (AI). Lignes rouges représentent les axones bout. (C) mesure et normalisation des régénératives des axones. Echelle = 20 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Discussion
Quand traverse le paramétrage de la mouche, le nombre de femelles et mâles utilisés peut varier selon les génotypes et le nombre de larves nécessaires pour les expériences spécifiques. Pour les mouches WT, Croix typique utilise 10 femelles et 5 mâles. La fenêtre de la collection peut être réduite, selon la précision de l’âge des larves. Par exemple, une période de 2 h collection donneront des larves d’une population plus homogène. Dans ce cas, à l’aide de femelles de 20 ou plus vierges à mettre en place les croix aidera produisent des œufs suffisamment. Le rendement dès le premier jour est généralement rare, donc recueillir la plaque avec les œufs deux jours ou plus après la mise en place de la chambre. Lorsque davantage de culture la plaque avec les œufs, il est recommandé de faire tremper le tissu dans le plat de Pétri de 60 mm en solution acide propionique 0,5 % au lieu de l’eau, qui sera non seulement aider à maintenir l’humidité dans le plat mais aussi d’éviter la croissance de moisissures.
Lorsque vous configurez les périphériques pour l’anesthésie de larves et de montage, s’assurer d’utiliser un plat en verre, parce que l’éther va fondre à travers des matières plastiques. Le verre plat est logé dans un plastique de 15 cm boîte de Pétri dans le cas d’une fuite. Le papier de soie permet de préserver l’éther dans le plat pour que la larve peut effectivement être assommée par la vapeur d’éther. À l’aide de laisser tomber les bouteilles pour stocker des aliquotes d’éther et en ajoutant éther gouttelettes avec le compte-gouttes en verre ambre est recommandé. Reconstituer l’éther et gardez toujours une couche d’éther liquide au fond du plat pour un effet optimal.
Au moment de l’exposition de l’éther est critique : sous anesthésie conduira à la reprise de la larve au milieu de la séance d’imagerie et des images tremblantes ; un surdosage de l’anesthésie, ou un contact direct avec le liquide de l’éther, provoquera la létalité. Pour maximiser la survie et le taux de réussite, notre règle de base est la suivante : PNS blessure/imagerie, sortez la larve dès que sa queue s’arrête tics ; pour le CNS, attendre que la larve entière devient immobile, en particulier les segments de tête. Même un léger mouvement interférera avec la blessure et l’imagerie des axones VNC. Larves âgées ont tendance à prendre plus de temps pour l’assommer. Il faut en général moins de 2 min pour les larves de moins de 72 h AEL et 2-5 min pour les larves de plus de 72 h AEL.
Lorsque vous configurez l’intensité du laser deux photons pour blessure, la valeur est déterminée par le signal de fluorescence de tissus obtenu à partir de l’analyse de « Live ». La blessure est introduite par un balayage « Continu » comme à l’étape 3.3. L’augmentation induite par la blessure de fluorescence est due à l’autofluorescence au site de la lésion et sert comme un bon indicateur de la rupture de la neurites. En règle générale, il faut s 1-10 de voir la pointe de la fluorescence. Il est essentiel de contrôler le temps de scan une fois que la fluorescence est élevée. Des blessures PNS, il est essentiel d’arrêter immédiatement la numérisation. Une exposition prolongée sera agrandir le site de la lésion et d’endommager les tissus voisins. Toutefois, cela offre la possibilité d’ajuster les temps de numérisation et de manipuler la gravité de la blessure. Une blessure réussie doit avoir un diamètre de taille de lésion plus petits que 3-4 μm. Des blessures VNC d’axon, les axones VNC sont incorporés beaucoup plus profond par rapport à la da neurone PNS axones/dendrites, qui sont juste sous la peau et donc nécessitent plus laser intensité. Habituellement, nous quittons le scan sur pendant quelques secondes de plus. C’est pour s’assurer que le paquet entier axon est rompu. Si le rayon des sites lésion n’est plus de la moitié de la largeur du faisceau commissure, ces sites de blessures sont comptés aussi infructueuses. Ces larves ont un faible taux de survie et ne figurera pas dans l’analyse.
Pour la régénération axonale, la capacité de régénération est semblable dans les stades larvaires. Mais pour la régénération de dendrite après une coupe simple dendrite, le potentiel de régénération est réduit après 72 h AEL19. Ainsi, les lésions axonales sont généralement effectuée à 48 h - 72 h AEL et blessures de dendrite 48 h AEL, avec l’évaluation de la régénération à 120h AEL. Les larves de 24h AEL peuvent également être utilisés, mais ils nécessitent un traitement plus attentionné, compte tenu de leur petite taille. Les larves se nymphosent après 120 h AEL, rendant plus difficile de l’imagerie. Par conséquent, notre point de terminaison est généralement 120 h AEL.
Quelle est la corrélation entre la dégénérescence et régénération ? Blessure PNS, après 24 h AI, normalement l’axon/dendrite distale en WT a complété la dégénérescence wallérienne tandis que régénération n’a pas commencé à ce moment. Par conséquent, nous pensons que chez les larves de WT, dégénérescence des neurites coupée n’a un impact très limité sur la régénération, voire pas du tout. Des blessures VNC, axones à 8 h AI ont déjà commencé à dégénérer, et après 24 h AI, la régénération axonale est observée alors que les débris axon pourraient encore être trouvés autour des sites de blessures. Les débris ne semblent pas bloquer la régénération. Toutefois, il est possible que dans certaines circonstances, il peut y avoir un chevauchement ou même une Diaphonie entre dégénérescence et régénération. En fait, il a été signalé que chez des souris âgées, dégagement de débris après des dommages de nerf périphérique est plus lente que celle chez les jeunes animaux. En même temps, réinnervation plus lente de la jonction neuromusculaire a été observée, ce qui peut être attribué pour le plus grand nombre d’obstacles régénération axones rencontre chez les vieux animaux. Étonnamment, cependant, les axones provenant d’animaux âgés régénèrent rapidement et reinnervate sites de jonction neuromusculaire efficacement quand ne pas confrontés avec débris32. Ceci suggère que faciliter le dégagement de débris pourrait être une stratégie possible pour favoriser la régénération.
Par rapport aux autres modèles de la neurogenèse, le modèle de blessure du neurone sensitif mouche présente des avantages uniques. Modèles murins habituellement durer des semaines ou mois pour effectuer et ne conviennent pas pour la réalisation des écrans génétiques à grande échelle ; C. elegans a seulement un système nerveux primitif qui ne peuvent pas étroitement récapituler les barrières de la régénération dans le SNC chez les mammifères ; différent des mammifères, les axones de CNS zebrafish régénèrent robuste. Le cycle de vie rapid de mouches, la polyvalence de la mouche de la génétique, l’accessibilité et structuration stéréotypée des axones/dendrites des neurones sensoriels mouches et les propriétés caractéristiques de la régénération des neurones sensoriels mouches – sous-type spécifique régénération dans la PNS et régénération limitée dans le système nerveux central – font de neurones dopaminergiques Drosophila un modèle attrayant pour étudier la neurogenèse. En outre, des études récentes de cellules ganglionnaires rétiniennes de souris concassée (CGR) suggèrent également que les sous-types neuronales portent compétence régénération distinctes ; certains sous-types RGC peuvent se régénérer, alors que d’autres dans le faisceau de nerfs apparemment homogène ne parviennent pas à repousser les33. Cette importante découverte suggère que des stratégies spécifiques au type neuronales devraient être exploitées pour favoriser la régénération et la récupération fonctionnelle et plaide énergiquement que l’induction des lésions axonales et analyse de régénération subséquente doit être effectuée dans un manière de sous-type spécifique neuronale. Par ailleurs, les déterminants moléculaires et cellulaires pour ce type de régénération-spécificité restent largement inconnu19,33. Par conséquent, le modèle de lésion du neurone da offre le paradigme idéal pour s’attaquer à ces questions.
Comparativement à la régénération axonale, études qui portent sur la régénération de la dendrite sont beaucoup plus rares. Blessure de dendrite se produit, comme dans le traumatisme crânien, accident vasculaire cérébral et de nombreuses formes de neurodégénérescence, pourtant ne sait presque rien pouvoir des dendrites réparer et réformer les connexions neuronales. Le modèle de lésion du neurone da offre à nouveau un système très accessible qui affiche la répétition stéréotypée, la spécificité de la classe et temporelle règlement19, afin d’explorer cette voie.
Il est également important de mentionner que bien qu’il soit possible de blesser un seul axone étiqueté dans le PNS, le préjudice moyen est effectué dans les résultats de la CNS dans la lésion d’un faisceau d’axones. Si vous le souhaitez, MARCM34 ou l’approche de35 clone FLP-out peut-être servir d’étiqueter les axones unique dans le système nerveux central. En outre, lorsque les cellules gliales sont simultanément étiquetés avec RDRF (Repo-Gal4, SAMU-RDRF), l’accumulation des procédés de cellules gliales est observée plus précisément sur le site de la lésion. En outre, l’expression de Ptp99A, l’homologue de mouche de chondroïtine sulfate protéoglycane (CSPG) phosphacan / PTPRZ1, est régulée sur le site de la lésion. Ptp99A Co se localise avec les cellules gliales et entoure le site de la lésion, formant une structure d’anneau comme semblable à ce qui a été signalé pour les cicatrices astroglials mammifères19,36,37. En conclusion, le modèle de blessure du neurone da, lorsqu’il est combiné avec d’autres types de cellules comme les cellules gliales ou des cellules immunitaires, des marqueurs permettra in vivo la surveillance en temps réel des interactions multicellulaires entre un neurone blessé et ses environs environnement.
Bien que ce modèle de lésion du neurone sensitif les larves de mouches nous offre des possibilités de trouver des régulateurs neurorégénération potentiels dans le PNS et CNS, il a encore plusieurs limitations. Tout d’abord, il n’est pas du haut débit au stade actuel. En général, 5-6 génotypes pourraient subir par une seule personne en une semaine. Il doit être optimisé afin d’effectuer un écran non biaisé. Deuxièmement, comme l’anesthésie de l’éther sur larves peut durer de plusieurs min à pas plus de 20 min, il n’est pas optimal pour l’imagerie à long terme. Ainsi, des moments spécifiques qui sont représentatifs de la dégénérescence axonale et régénération respectivement sont choisis pour l’imagerie. Troisièmement, même si ce protocole met en place un moyen de blesser des axones avec précision, la possibilité de dommages aux tissus environnants ne peut pas être totalement écartée. Dans le VNC, ces tissus peuvent être des cellules gliales, les axones et les dendrites des autres neurones. Pour minimiser cette restriction éventuelle, nous réduire au minimum les sites de blessures, s’appliquent le plus bas possible de puissance de laser et en parallèle à la fois contrôler et expérimenter des groupes, exécutez les procédures. Quatrièmement, dans le processus de mise en place le paradigme de blessure de CNS, sites de différentes blessures ont été testés, y compris les sites près de terminus du axone que nous avons choisi d’utiliser et la point d’entrée des axones dans le VNC. Les points d’entrée se sont révélés pour être plus difficile à blesser, parce qu’ils sont plus profondément dans le tissu et plus susceptibles de se déplacer. Ainsi, pour ces raisons pratiques, nous optons pour la méthode actuelle, qui est bon pour des expériences à grande échelle, plus cohérentes et mieux contrôlée. Un seul souci potentiel, comme indiqué ci-dessus, est la possibilité de blesser les tissus voisins, tels que les éléments postsynaptiques et les cellules gliales. En revanche, c’est un remarquable question comment les tissus environnants endommagés influencent la dégénérescence et la régénération des axones. Par exemple, comment la cicatrice gliale affecte la régénération axonale. Il y a une autre raison pourquoi notre modèle de blessures peut-être sont très proches des modèles de lésion chez les mammifères, dont les axones et les tissus autour des sites de lésions sont habituellement blessés simultanément.
Blessure de laser suivi par microscopie Time-lapse est une analyse sensible pour l’étude de la régénération axonale/dendrite. Cependant, une des préoccupations principales de ce test sont le coût perçu, qui s’étend de < 10K $ pour les lasers à impulsions à l’état solide bas de gamme, $25-100K pour les lasers femtoseconde à >$ 100K pour les lasers de deux photons. Il y a plusieurs bonnes discussions sur les systèmes laser différentes utilisées pour axotomie29,38,39,40,41,42,43, 44 , 45. pour résumer, les lasers conventionnels sont optimales pour axones coupe au sein de 30 à 50 µm de la surface. Il y aura plus de dégâts collatéraux avec les lasers deuxième nano et pico, comparé avec le laser femtoseconde, autant que la profondeur de la zone cible augmente de45. Pour blesser les axones dans le VNC, dont la profondeur est généralement environ 50-100 µm, il est essentiel de minimiser les dommages aux tissus. Dans ce cas, le deux photons laser est idéal, qui porte la puissance du laser pour le plan focal, réduisant les dégâts collatéraux de tissus sans compromettre la pénétration tissulaire. En conclusion, le système de deux photons est coûteux, mais offre la meilleure préservation précision et tissus. Toutefois, si seulement les axones PNS sont la cible d’une axotomie, lasers à impulsions conventionnel peuvent être une alternative plus abordable.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous remercions Jessica Goldshteyn pour le support technique. Travail dans le laboratoire de la chanson est financé par la subvention du NIH R00NS088211 et l’intellectuel et Developmental Disabilities Research Center (IDDRC) nouveau programme Development Award.
matériels
| Name | Company | Catalog Number | Comments |
| Diethyl ether, ACS reagent, anhydrous | Acros Organics | AC615080010 | |
| Halocarbon 27 Oil | Genesee Scientific | 59-133 | |
| Phosphate buffered saline (PBS), 20x Concentrate, pH 7.5, supplier # E703-1L | VWR | 97062-948 | |
| Agar powder, Alfa Aesar, 500GM | VWR | AAA10752-36 | |
| Grape juice | Welch’s | ||
| Ethanol 95% (Reagent Alcohol 95%) | VWR | 64-17-5 | |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Propionic Acid | J.T.Baker | U33007 | |
| Cover Glasses: Rectangles | Fisher Scientific | 12-544-D | 50 mm X 22 mm |
| Zeiss LSM 880 laser scanning microscope | Zeiss | ||
| Zen software | Zeiss | ||
| Chameleon Ultra II | Coherent |
Références
- Yakura, J. S. Recovery following spinal cord injury. , (1996).
- Harel, N. Y., Strittmatter, S. M. Can regenerating axons recapitulate developmental guidance during recovery from spinal cord injury? Nature reviews. Neuroscience. 7, 603-616 (2006).
- Jurewicz, A., Matysiak, M., Raine, C. S., Selmaj, K. Soluble Nogo-A, an inhibitor of axonal regeneration, as a biomarker for multiple sclerosis. Neurology. 68, 283-287 (2007).
- Yiu, G., He, Z. Glial inhibition of CNS axon regeneration. Nat Rev Neurosci. 7, 617-627 (2006).
- Sun, F., He, Z. Neuronal intrinsic barriers for axon regeneration in the adult CNS. Curr Opin Neurobiol. , (2010).
- Liu, B. P., Cafferty, W. B., Budel, S. O., Strittmatter, S. M. Extracellular regulators of axonal growth in the adult central nervous system. Philos Trans R Soc Lond B Biol Sci. 361, 1593-1610 (2006).
- Liu, K., Tedeschi, A., Park, K. K., He, Z. Neuronal intrinsic mechanisms of axon regeneration. Annu Rev Neurosci. 34, 131-152 (2011).
- Schwab, M. E., Strittmatter, S. M. Nogo limits neural plasticity and recovery from injury. Curr Opin Neurobiol. 27, 53-60 (2014).
- He, Z., Jin, Y. Intrinsic Control of Axon Regeneration. Neuron. 90, 437-451 (2016).
- Park, K. K., et al. Promoting axon regeneration in the adult CNS by modulation of the PTEN/mTOR pathway. Science. 322, 963-966 (2008).
- Geoffroy, C. G., Hilton, B. J., Tetzlaff, W., Zheng, B. Evidence for an Age-Dependent Decline in Axon Regeneration in the Adult Mammalian Central Nervous System. Cell Rep. 15, 238-246 (2016).
- Geoffroy, C. G., et al. Effects of PTEN and Nogo Codeletion on Corticospinal Axon Sprouting and Regeneration in Mice. J Neurosci. 35, 6413-6428 (2015).
- Jin, D., et al. Restoration of skilled locomotion by sprouting corticospinal axons induced by co-deletion of PTEN and SOCS3. Nat Commun. 6, 8074(2015).
- Wang, X., et al. Axonal regeneration induced by blockade of glial inhibitors coupled with activation of intrinsic neuronal growth pathways. Exp Neurol. 237, 55-69 (2012).
- Fang, Y., Bonini, N. M. Axon degeneration and regeneration: insights from Drosophila models of nerve injury. Annual review of cell and developmental biology. 28, 575-597 (2012).
- Venken, K. J., Simpson, J. H., Bellen, H. J. Genetic manipulation of genes and cells in the nervous system of the fruit fly. Neuron. 72, 202-230 (2011).
- Leyssen, M., et al. Amyloid precursor protein promotes post-developmental neurite arborization in the Drosophila brain. The EMBO journal. 24, 2944-2955 (2005).
- MacDonald, J. M., et al. The Drosophila cell corpse engulfment receptor Draper mediates glial clearance of severed axons. Neuron. 50, 869-881 (2006).
- Song, Y., et al. Regeneration of Drosophila sensory neuron axons and dendrites is regulated by the Akt pathway involving Pten and microRNA bantam. Genes Dev. 26, 1612-1625 (2012).
- Song, Y., et al. Regulation of axon regeneration by the RNA repair and splicing pathway. Nat Neurosci. 18, 817-825 (2015).
- Kato, K., Forero, M. G., Fenton, J. C., Hidalgo, A. The glial regenerative response to central nervous system injury is enabled by pros-notch and pros-NFkappaB feedback. PLoS Biol. 9, e1001133(2011).
- Fang, Y., Soares, L., Teng, X., Geary, M., Bonini, N. M. A novel Drosophila model of nerve injury reveals an essential role of Nmnat in maintaining axonal integrity. Curr Biol. 22, 590-595 (2012).
- Xiong, X., et al. Protein turnover of the Wallenda/DLK kinase regulates a retrograde response to axonal injury. J Cell Biol. 191, 211-223 (2010).
- Brace, E. J., DiAntonio, A. Models of axon regeneration in Drosophila. Exp Neurol. 287, 310-317 (2017).
- Hao, Y., Collins, C. Intrinsic mechanisms for axon regeneration: insights from injured axons in Drosophila. Curr Opin Genet Dev. 44, 84-91 (2017).
- Galbraith, J. A., Terasaki, M. Controlled damage in thick specimens by multiphoton excitation. Mol Biol Cell. 14, 1808-1817 (2003).
- Sugimura, K., et al. Distinct developmental modes and lesion-induced reactions of dendrites of two classes of Drosophila sensory neurons. J Neurosci. 23, 3752-3760 (2003).
- Yanik, M. F., et al. Neurosurgery: functional regeneration after laser axotomy. Nature. 432, 822(2004).
- Wu, Z., et al. Caenorhabditis elegans neuronal regeneration is influenced by life stage, ephrin signaling, and synaptic branching. Proc Natl Acad Sci U S A. 104, 15132-15137 (2007).
- Stone, M. C., Nguyen, M. M., Tao, J., Allender, D. L., Rolls, M. M. Global up-regulation of microtubule dynamics and polarity reversal during regeneration of an axon from a dendrite. Mol Biol Cell. 21, 767-777 (2010).
- Grueber, W. B., Jan, L. Y., Jan, Y. N. Tiling of the Drosophila epidermis by multidendritic sensory neurons. Development. 129, 2867-2878 (2002).
- Kang, H., Lichtman, J. W. Motor axon regeneration and muscle reinnervation in young adult and aged animals. J Neurosci. 33, 19480-19491 (2013).
- Duan, X., et al. Subtype-specific regeneration of retinal ganglion cells following axotomy: effects of osteopontin and mTOR signaling. Neuron. 85, 1244-1256 (2015).
- Lee, T., Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron. 22, 451-461 (1999).
- Grueber, W. B., et al. Projections of Drosophila multidendritic neurons in the central nervous system: links with peripheral dendrite morphology. Development. 134, 55-64 (2007).
- Buss, A., et al. NG2 and phosphacan are present in the astroglial scar after human traumatic spinal cord injury. BMC Neurol. 9, 32(2009).
- McKeon, R. J., Jurynec, M. J., Buck, C. R. The chondroitin sulfate proteoglycans neurocan and phosphacan are expressed by reactive astrocytes in the chronic CNS glial scar. J Neurosci. 19, 10778-10788 (1999).
- Raabe, I., Vogel, S. K., Peychl, J., Tolic-Norrelykke, I. M. Intracellular nanosurgery and cell enucleation using a picosecond laser. J Microsc. 234, 1-8 (2009).
- Hutson, M. S., Ma, X. Plasma and cavitation dynamics during pulsed laser microsurgery in vivo. Phys Rev Lett. 99, 158104(2007).
- Venugopalan, V., Guerra, A. 3rd, Nahen, K., Vogel, A. Role of laser-induced plasma formation in pulsed cellular microsurgery and micromanipulation. Phys Rev Lett. 88, 078103(2002).
- Bourgeois, F., Ben-Yakar, A. Femtosecond laser nanoaxotomy properties and their effect on axonal recovery in C. elegans. Opt Express. 16, 5963(2008).
- O'Brien, G. S., et al. Two-photon axotomy and time-lapse confocal imaging in live zebrafish embryos. J Vis Exp. , (2009).
- Tsai, P. S., et al. Plasma-mediated ablation: an optical tool for submicrometer surgery on neuronal and vascular systems. Curr Opin Biotechnol. 20, 90-99 (2009).
- Chung, S. H., Clark, D. A., Gabel, C. V., Mazur, E., Samuel, A. D. The role of the AFD neuron in C. elegans thermotaxis analyzed using femtosecond laser ablation. BMC Neurosci. 7, 30(2006).
- Williams, W., Nix, P., Bastiani, M. Constructing a low-budget laser axotomy system to study axon regeneration in C. elegans. J Vis Exp. , (2011).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.