
Method Article
記録と電極アレイに結合された齧歯動物の脳スライスにおけるてんかん活動の変調
要約
微小電極アレイを用いたげっ歯類脳スライス 4 アミノピリジン誘発てんかん活動の記録と電気変調を実行する方法を示します。カスタム記録室は、長期の実験的セッションを通して組織実行可能性を維持します。電極マッピングをライブ、刺激的なペアの選択は、カスタム グラフィカル ユーザー インターフェイスによって実行されます。
要約
側頭葉てんかん (TLE) は、最も一般的な部分複雑なてんかん症候群と薬に応答性が最小です。薬理学的治療の障害や、脳神経はお勧めしませんが、脳深部刺激療法 (DBS) は有望なアプローチです。微小電極アレイ (Mea) と結合した急性期脳スライスは、電気刺激による神経ネットワークの相互作用とその変調を研究するための貴重なツールを表します。従来の細胞外記録法と比較すると彼らは観測点と既知の電極間距離、伝搬経路とスピード, 電気生理学の勉強をするのより多くの利点を提供します。通知します。ただし、組織酸素化が MEA 記録、減少した信号対雑音比と高い振動実験温度を犠牲にして来る高灌流率を必要とする時に大きく損なわれます。電気刺激をさらに難しく長期記録/刺激の新紀元を追求する脳組織に重点を置きます。また、脳スライスの電気変調は、電極マッピング簡単かつ迅速にライブ中に実行する実験を必要とする脳スライス内の特定構造/経路を対象とする必要があります。ここでは、4-アミノピリジン (4AP) の記録と電気変調を実行する方法を示します-プロポフォ平面測定の使用齧歯動物脳スライスを誘発。マウスから得られた脳組織ラット脳組織よりも優れているし、MEA の実験に適していますこのようにことを示す.このプロトコルは、世代と忠実に従来フィールド電位計測による電気生理学的特徴を再現、数時間持続する、長持ちする安定したてんかんパターンのメンテナンス維持保証します。電気刺激による長期新紀元。実験を通して組織の生存率は、層流と低 (1 mL/分) でもクイック ソリューション exchange 灌流率を可能にする少量のカスタム記録室の使用のおかげで達成されます。リアルタイムの監視と刺激電極の選択クイック MEA のマッピングは、カスタム グラフィック ユーザー インターフェイス (GUI) によって実行されます。
概要
てんかんは、脳の1; の自由な活動を引き起こす生命を脅かす進行性の障害それは病気および重要な社会的な汚名2,3の最も高い負担の間で運ぶ。TLE は最も頻繁に症候群 (40%) と最もよく (~ 30%) 抗てんかん薬4に耐性。てんかん組織の外科的切除は、患者さんの状態を改善するかもしれません、それすべての患者で現実的ではないと5完全に発作のない生活を保証しないかもしれない。薬物治療や脳神経外科が最適でない場合、電気 DBS によるてんかんの辺縁系ネットワークの変調、有望なアプローチです。
齧歯動物の脳のスライスは、彼らを少なくとも一部、元のアーキテクチャおよび脳の接続に保存電気生理学技術の in vitroによる健康と病気6どのように神経ネットワーク機能を研究するための貴重なツール利益 (率 ROI) の地域。特に、水平結合された海馬内嗅皮質 (海馬-EC) スライス TLE に関わる重要な神経ネットワークを構成して、したがって、定期的に体外TLE 研究7で採用されています。
人工髄液 (アプライド)、カリウム8,9,10を増加させながらマグネシウムの減少などのイオン組成を変更することによって急性発作のような活動脳スライスで誘導することがありますか抑制性の gaba 作動性活性をブロックするなどの薬理学的操作による (11包括的なレビューを参照してください)。ただし、これらのモデルは、興奮と抑制; のアンバランスの変質に基づいていますしたがって、彼らは相互作用と発作原性に興奮性と抑制性ネットワークの協調的貢献を勉強とは認めない。痙攣薬 4AP と脳スライスの持続灌流がそのまま全体的にシナプスの活性を保ちながら急性発作原性を研究するように、興奮性と抑制性の神経伝達を高める12。
測定できる空間的な制約ことができる電極の数を制限、従来の細胞外フィールド電位計測と比較して観測点の大きい数から神経ネットワークによって生成された電気活動を記録脳スライス面に収容されます。さらに、MEA チップの既知の電極間距離をトレース伝播に非常に便利です追加の利点を提供し、記録された信号の進行速度を評価します。培養神経細胞13,14からの録音の最初の着想、MEAs 今も使用目的は齧歯動物15と人間16から得られた急性脳スライスの電気生理学的特徴を特徴付けます。つまり、てんかん研究のコンテキスト、Mea は発作原性16,17,18の中核神経ネットワークの相互作用を特定するための貴重なツールを表します。
しかし、MEA 記録は本質的に取得または長時間 (数時間) 実験的プロトコル全体の安定したてんかんパターンを維持する技術的な課題を運ぶ。まず、組織酸素化がない大きい内になりぐるぐる水没型記録室19,20市販 MEAs の典型的な貧しい信号対雑音比と温度不安定性に影響を与える可能性があります。(たとえばテクニカル ノートの参照暖房と灌流装置21) 脳スライスへの酸素供給を改善するために使用する高灌流率 (6-10 mL/分) 録音の品質。水没型記録室19; を使用する場合、てんかん放電などの高周波成分を備えた第二に、再発の放電が観察されるほとんどこれは、特に急性てんかんパターンが化学的に誘導され、ephaptic メカニズムが含まれます 4AP モデル22のためのケースである (11の包括的な概観を参照)。これらの制限を克服するために研究者によっていくつかの方法が提案されています。たとえば、脳スライス厚 (≤300 μ M) とエリア17,18を削減しながら灌流率19,20を増加する組織に十分な酸素供給を達成するために重要な要因であります。さらに、改善脳スライスの生存率は、両方の側面の18から脳組織を灌を許可する穿孔 Mea (pMEAs) を使用しても実現できます。
説明のアプローチは、脳スライスから MEA 記録の可能性を改善している大幅に対し彼らがテストされていないに対して長時間 (数時間)、後者を表す重要な記録と電気刺激セッション脳組織のストレス。長期にわたるレコーディング セッションは、短期的な測定によるマスクを解除することはできませんてんかんパターンの特定の機能の時間の進化を研究する必要があります。DBS 研究においては、長期にわたる実験的プロトコルを評価し、同じ脳スライス内のいくつかの刺激パラダイムの効果を比較する必要があります。
自発の電気活動だけが投資収益率を基準電極のマッピングを記録する必要がある場合、通常データ分析; 中には事後、すなわち、行われます代わりに、誘発または電気ニューロモデュレーション パラダイムの研究は、刺激が、実験中に迅速かつ簡単なライブ電極マッピングの必要性を口述特定 ROI(s) に配信されることが必要です。
ここでは、我々 は大人の齧歯動物の脳切片で安定した 4AP 誘発てんかんパターンの誘導と維持は、簡単な実験プロトコルを示しています。観測活動は、従来の細胞電気生理学手法の特徴としてこのモデルの電気生理学的特徴を忠実に再現します。それは数時間持続して繰り返し長時間新紀元の持続的な電気刺激を長持ち。維持し、脳スライスをインキュベート室組み立てることができます簡単に標準的な実験室の供給 (図 1 a) を使用して最適なソリューション為替レートと流 (図 1 b) を可能にするカスタム録音室ができるに対し商業ソースから取得する、または手頃な価格の 3 D 印刷技術を使用しています。リアルタイム監視、および刺激的な電極の選択 ROI(s) の迅速なマッピングは、 mapMEA、自由にリクエストを名前付きカスタム ユーザーフレンドリーな GUI で可能です。

図 1: このプロトコル用のカスタム装置。(A) 回復済み温暖化、4AP で前培養室は、ビーカー、シャーレを使用して組み立てている保持。ペトリ皿はブレーカーより直径の小さい、シリンジのプランジャーによって行われています。ペトリ皿の底は、ペトリ皿の縁にシアノアクリ レート接着 (ソフト ストッキング) からナイロン メッシュに置き換えられます。酸素は、ペトリ皿の壁とビーカーの間挿入曲がった脊髄針 (22 G) を介して提供されます。泡は、ビーカーと決してリーチ脳スライス変位を避けるためにナイロン メッシュの側から浮上する必要があります。シャーレの上部は、アプライドの蒸発を避けるために、酸素飽和度を維持、蓋として使用できます。(B) カスタム記録室。: 暖房カニューレ及び参照電極に合わせて入口貯水池。うち: 吸引針に合わせて出口貯水池。rec: レコーディング室。 (C) ガラス パスツール ピペット、火研磨によってカール組織を処理し、記録室内での位置を調整するために使用します。 (D) カスタム スライス ホールド アンカー。(E) 記録室の最終組立は、MEA チップに搭載。脳スライスは、アンカーによって行われての下にかかっています。赤の矢印は赤マル印参照電極、インレット タンクのアプライドで水没した飽和 KCl ペレットに対し暖房カニューレをカバー PTFE チューブ。青の矢印の出口貯水池の吸引針。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
プロトコル
ここで説明するすべての方法は、EU 指令 2010/63/EC 動物の福祉に準拠してイタリアの保健 (承認 (名) 860/2015-PR) 省によって承認されています。
1. 商工会議所 MEA/カスタム アセンブリの準備
注意: 起動実験前に 2 日間です。リングなし MEA を使用 (材料表を参照してください)。
-
きれいに MEA (期間: 35 ~ 40 分)。
- 酵素クリーナー パウダーを温かい蒸留水に溶解します。ブラシの MEA 表面洗浄暖かいし、MEA を浸して洗浄剤 3 h、少なくとも「暖かい。
- 蒸留水で十分に MEA を洗うし、糸くずのない傷のティッシュを使用してそれを乾燥します。
-
録音室で MEA を組み立てる (期間: 10 分 + 一晩)。
- 録音室の底面に弾性シール材を均等に分散します。泡がないことを確認します。
- ピンセットの助けを借りて、MEA に録音室をマウントし、穏やかな圧力を適用します。任意の気泡がある場合は、円で商工会議所を移動できますわずかすべての泡が見えなくなるまで指で穏やかな圧力を適用している間。録音室の外側の境界線の周りにすべての方法シーラント層を広げます。
注:重要な!シーラントと MEA の接触をカバーしていません。 - MEA/商工会議所アセンブリ 15 cm シャーレ内に置きます。場所シャーレの 5 mL または 10 mL のビーカーは、MEA の近くに蒸留水を満たした。湿潤環境を維持するためにすべてをカバーします。一晩常温で治療をしましょう。
注: プロトコルはここで一時停止することができます。
-
コート親水性させる MEA (期間: 5-10 分 + 一晩)。
- 15 cm のシャーレに MEA を配置します。ポリ-D-リジンの 50 μ L を注ぎ MEA 記録領域に。ペトリ皿を閉じ、フィルム、シールで封印、4 ° C で一晩保管
- 蒸留水を使用して徹底的に MEA をリンスし、蒸留水で 4 ° C で MEA を格納します。
注: プロトコルはここで一時停止することができます。コーティング測定は格納され、いくつかの実験の再利用があります。定期的にクリーニングと再コーティングの MEA ティッシュの残骸を削除し、信号対雑音比を向上させることができます。
2. ストック溶液の調製
注: 起動実験前に 1 日 (期間: ~ 2 h)。在庫ソリューションは、実験の日に実験の高速化を支援します。彼らは、事前に準備して格納できます。集中率と組成の表 1を参照してください。最終的なソリューションの構成は表 2を参照してください。
- 室温で蒸留水中の化学物質を溶解します。フィルターのボトルに原液をフィルター (フィルター径: 0.22 μ m)。
- 4 ° C でストア (推奨最大保存時間は、表 1 を参照)。
注: プロトコルはここで一時停止することができます。
3. 寒天培地の調製
注: 起動実験前に 1 日 (期間: ~ 1 h)。
- 250 mL の蒸留水を 500 mL のビーカーに注ぐし、350 rpm で攪拌を開始します。寒天 5 g を追加し、それを完全に溶解するまで待機します。
- 熱 250-300 ° C でソリューションソリューションが 2.5 %w/v 寒天液を屈する 〜 200 mL になるまで蒸発水をしましょう。
注: 温度が十分に高い水が泡立ち無し蒸発させるはずです。 - Cm 高壁 ~1.5 の 200 mL プラスチック正方形のボックスの上に寒天液を注ぎ、冷まします室温で固体になるまで。
- ボックスをカバーし、4 ° C で保管寒天培地のブロックは、数週間のために良いです。
注: プロトコルはここで一時停止することができます。
4. 凍結面の準備
注: は、実験前に 1 日を開始します。
- 0.5 - 1 cm 縁下まで蒸留水でフラット底ステンレス ボウルを埋めます。
- 一晩-20 ° C で保存します。
5. 神経切断アプライドの準備
注: 起動実験前に 1 日または実験の同日 (表 1と表 2を参照) (期間: 30 分)。
- 500 mL ビーカーに蒸留水 200 mL を注ぎ、350 rpm で磁性攪拌器を使用して攪拌を開始します。
- ストック B の 50 mL と 50 mL の在庫 C の追加 (表 1参照)。
- 1 mM L-アスコルビン酸、D-グルコース 10 mM 208 mM ショ糖 3 mM ピルビン酸を追加します。
注: ピルビン酸の実際の量は密度や純度に依存し、バッチによって異なる場合があります。常に新しいバッチを開くときに必要量を計算します。 - フィルム シール カバーし、30 分間攪拌させます。
- 500 mL のメスフラスコに溶液を移し、フラスコに蒸留水を追加します。
- フィルム シールと容積測定フラスコを密封し、優しくの反転 3 - 5 回ソリューションが均等にクリアされるまで。
- キャップをガラス瓶の中に切断アプライドを転送します。
- 4 ° C まで冷却するため、切削アプライド-80 ° C、20-30 分または 4 ° C で一晩を格納します。
注: プロトコルを一時停止できるここで一晩冷却が好ましい場合。それ以外の場合所要時間冷却切削アプライドを使用して手順 6 を追求する可能性があります。
6. 開催の準備アプライド
注: 準備ソリューション新鮮な実験の日 (期間: 5-10 分)。アプライドはスライスのプロシージャの間に切削アプライドから脳スライスをすすぎに使用する保持 (手順 8.16 参照) と脳スライスの長期的なストレージおよび温暖化済み。脳スライスをすすぎにアプライドの 100 mL で十分です。このプロトコルで使用される保持および事前地球温暖化の部屋は、カスタムメイドされそれぞれ 300 mL と 100 mL の量が含まれて (参考: 図 1 a)。この設定では、アプライドの 500 mLで十分です。商業ソースから得られたチャンバー ケース開催準備するアプライドの全体量を調整する必要があります別のボリュームがあります。
- 500 mL のメスフラスコに蒸留水 200 mL を注ぐ。
- 1 mM L アスコルビン酸株式 M、6.5 mL、50 mL のストック B の株式 A の 50 mL を追加します。振とう L-アスコルビン酸を完全に溶解するまで、回転の動きを使用して。
- 最終巻に到達、フィルム、シールとシールの容積測定フラスコに蒸留水を追加し、そっとの反転 3 - 5 回ソリューションが均等にクリアされるまで。持株アプライド組成は表 2を参照してください。
7. 実験ベンチの準備
注: これは 15 分プラス 30 分暖かい浴室の安定した温度を取得する必要があります。
-
記録ベンチ
- 温かいお風呂を開始、32 ° C に設定して、その温度を維持するためにそれをカバーします。
注: このプロトコルで使われる温かいお風呂は、カスタムメイド ハード プラスチック製のボックスと所望の温度で設定済み水槽サーモスタットを使用することができます。安定した温度は、室温から始まって ~ 30 分で達して可能性があります。前地球温暖化とインキュベーターのチャンバーは、プラスチック製のボックスの下部に座っている、ので水位はする必要があります浮動小数点から部屋を保つためにサーモスタットをまかなうに足るだけ。 - 開催と事前地球温暖化室 (参考: 図 1 a)、組み立てるし、それらにアプライドの持株を注ぐ。室温で保持チャンバーを維持し、事前地球温暖化室内の温かいお風呂を配置します。バブル 95% O25% CO2ガス混合ソリューションを開始し、反転パスツール ピペットを使用して任意の閉じ込められた気泡を削除します。覆われて保ちます。
- 温かいお風呂を開始、32 ° C に設定して、その温度を維持するためにそれをカバーします。
-
ベンチと vibratome スライス
- シアノアクリ レート系接着剤を使用して試料ディスクの中央に小さな寒天ブロックを接着します。
- 氷のバケツに 250 mL のビーカーを置くし、アプライドに食い込んでそれを注ぐ。
- 2 つの 100 mL ビーカーを取るし、それぞれにアプライドの 50 mL を注ぐ。室温でビーカーを残します。すすぎのビーカーです。
- バブル 95% O25% CO2ガス混合ソリューションを開始します。
- Vibratome にバッファー トレイをマウントし、砕いた氷とそれを囲みます。スライス処理中に低温を維持するために支援するために砕いた氷をいくつかエタノールを注ぐ。注ぐエタノール後必要に応じてより多くの氷を追加します。
注: 2 ° C の温度が最適です。 - バッファー トレイ内バブラーを配置し、組み立てる vibratome ブレード ブロックをマウントします。
注: クリアランス ~ 10 ° の角度でブレードがマウントされていることを確認します。 - ボリュームと蒸留水と塗りの 2/3 に 500 mL ビーカーに砕いた氷を設置します。
- 冷凍庫から冷凍のボールを取り、スライスのベンチに逆さまに配置、紙タオル、ろ紙ディスクでそれをカバーします。
-
麻酔ベンチ
- 氷で満たされたトレイに小さなボウルを配置し、アプライドをカットを注ぐ。95% O25% CO2ガス混合物でバブルを開始します。
8. 脳スライス標本とメンテナンス
注: このプロトコル利用大人 (4 - 6 週齢) の男性 CD1 マウスが他の系統 (例えばc57/bl617,18) を使用できます。我々 はまた後でと同じ年齢の男性の Sprague-dawley ラットから得られた脳スライスによって得られたマウス脳切片を用いて実験の出力を比較します。次の手順は、CA3 駆動高速間欠期のような活動が適切な海馬が拘束され、として海馬傍皮質に伝達できない脳スライス部分的切断の準備を参照してください。23で説明します。海馬-皮質の切断は、海馬 EC 脳スライス標本を用いた電気変調研究を追求する前提です。使用 vibratome は、材料表に記載されているモデルです。他のモデルは、別のプロシージャを必要があります。
- イソフルランの使用齧歯動物を麻酔 5%、カーボゲン ・ ガス混合物 2 L/分で麻酔誘導室に配信。
- 深麻酔下 (足への応答と足反射ピンチ)、23のように標準的な手順 1 min 以内で脳を抽出します。釣合い冷たい切削アプライドを含む小さなボウルに脳を置き、90-120 秒冷やします。
注: が長すぎるためチル脳をさせない、それがフリーズするそれ以外の場合。 - バッファー トレイに冷たい切断アプライドを注ぐ。フィルター紙の上に広がる切削アプライドは冷凍のボウルに配置されます。フィルター紙の上に脳を置き、小脳を取り外して、まっすぐ正面ポールを切る必要な脳組織ブロックを分離します。
- 曲がったへらの助けを借りて、カット前頭面寒天ブロックと vibratome 刃に直面して後頭極にして試料ディスク上に脳の背側を接着します。シアノアクリ レート系接着剤の薄層を使用します。すぐにバッファー トレイに試料ディスクを置き、固定します。
- 寒天ブロックまで組織横断 vibratome ブレードを移動するために断面範囲を調整します。脳組織ブロックとブレードのレベルに到達する試料ディスク高さを調整します。
- 海馬がはっきり表示されるまでティッシュ セクションを破棄 (通常 ~ 900 μ m)。400 μ m のスライス厚を設定し、組織切片を保持するスライスを開始します。各脳のセクションの 2 つの半球を分割し、2 つの脳のスライスを取得する不要な組織をトリムします。バッファー トレイはその後のセクションの中に高まって、アイスの蒸留水でバッファー トレイ拠点を埋める (7.2.7 の手順を参照してください)。
- 軽く最初の洗浄のビーカーで脳スライスを転送、パスツール ピペットを空にして脳スライスを 2 回目のすすぎのビーカーに転送反転パスツール ピペットを使用します。保持室に脳スライスを転送し、少なくとも 60 分回復させます。
注: 通常 6-8 脳スライス全体を取得することが可能です。プロトコルはここで一時停止する可能性があります。
9. 4 AP (4AP アプライド) を含むアプライドの準備
注: これは、準備のため 10 分プラス 20 分地球温暖化の必要があります。注意!4 AP は痙攣の薬、それは有毒であります。手袋をはめて処理し、こぼれます。
- 500 mL のメスフラスコに蒸留水 200 mL を注ぐ。
- 1 mM L アスコルビン酸株式 M、5 mL、50 mL のストック B の株式 A の 50 mL を追加します。振とう L-アスコルビン酸を完全に溶解するまで、回転の動きを使用して。
- 4AP 在庫の 500 μ L を追加します。
- 最終巻に到達、フィルム、シールとシールの容積測定フラスコに蒸留水を追加し、ソリューションが均等にクリアされるまでを優しくそれに 3-5 回を反転します。
- (参考: 図 1 a) 4AP インキュベーターのチャンバーを組み立てるし、4AP アプライドの ~ 80 mL を注ぐ。商工会議所をカバー、温かいお風呂にそれを転送し、95% O25% CO2ガスの混合物でバブリングを開始します。反転パスツール ピペットを使用して任意の閉じ込められた気泡を削除します。覆われて保ちます。
- 4AP アプライド温度は 30-32 ° C (~ 20 分) まで待機します。
注: プロトコル可能性がある場合に一時停止ここ脳スライスはまだ回復します。
10. 事前地球温暖化と 4AP のスライスの孵化 (期間: 90 分)
- 事前に地球温暖化のアプライド ・ 4AP アプライド温度が 30-32 ° c. であることを確認します。
- 逆ガラス パスツール ピペットを使用して事前地球温暖化室に 1 脳スライスを転送し、25-30 分間組織休ませて。4AP アプライドのインキュベーター室に脳スライスを転送し、60 分間安静にしましょう。
11. MEA セットアップの準備
注: は、録音の前に 15 分を開始します。
- 4AP アプライドの残りの量を 500 mL の三角フラスコに転送します。
- MEA アンプの上の棚に三角フラスコを置き、60 mL の注射器を連続的に給紙するソリューションを許可するようにチューブを使用します。三角フラスコと 1 mL/分の重力供給率を許可するように注射器の高さを調整します。
注: 適切な高さは、チューブ内径 (ID) に依存します。5/32 インチ ID のチューブ、30 cm あれば十分です。 - バブル 4AP アプライド エルレンマイヤー フラスコと 95% O25% CO2ガス混合物で注射器を起動します。
- そこまでビーカーに灌流管を通して聞かせて 4AP アプライド流れは内部; 空気がないです。ソリューション フローを停止します。
- MEA の基地で加熱要素をサーモスタットに接続します。MEA アンプ内部乾燥の MEA チップを置き、アンプ ヘッドを保護します。プラスチックのパスツール ピペットを使用して 4AP アプライドを入口と録音室の外側の貯水池に転送。
- マグネット ホルダーに暖房カニューレをセキュリティで保護し、記録室入口ポート内にその先端を配置MEA アンプ ヘッド; の磁気ストライプに磁気ホルダーを取り付けますカニューレ; に灌流管を接続します。カニューレをサーモスタットに接続します。
注: 加熱カニューレは、傾斜したポリテトラフルオロ エチレン (PTFE) チューブ記録室入口ポートに到達してカニューレの金属材料によるノイズを最小限に抑えることによってカバーされなければなりません。インレット タンクが十分に 4AP アプライドでいっぱいになる、灌流システムから落ちる水滴は表示しないでください。 - 貯水池内吸引針を置き、アプライド; に吸引針を水没によって否定的な圧力があることを確認一定の低周波吸引音を確認します。
注: 吸引針は脳スライス面の直上流に 4AP アプライドを許可するように置かれるべき。低騒音真空ポンプや真空ラインに使用できます。 - 1 mL/min の流速にでき始める灌流量制御を設定します。
注: 重力供給の灌流は、蠕動ポンプによって引き起こされるかもしれないノイズを除去します。蠕動ポンプが好まれる低雑音モデルは必須です。 - 4AP アプライド、カニューレを流れる、一度サーモスタットを入れます。37 ° C および MEA ベース録音チャンバ内 32-34 ° C の温度を達成するために 32 ° c に加熱カニューレを設定します。
注:注意!決して熱またはそれのソリューションなしカニューレが不可逆的破損があります。カニューレの固有温度オフセット設定値と記録室内暖房カニューレの先端では実際の値との間に記録温度 (すなわち32-34 ° C) よりも高い温度を設定します。流量、温度、環境、録音時の音量は部屋すべて影響記録ソリューションの温度です。11.9 の手順で報告された設定は、記述されているプロトコルおよび装置に最適です。常に熱電対を使用して実際の録音の温度をチェックし、必要に応じて設定を調整します。脳スライスの過熱を避けるために 34 ° C の上の MEA ベースを加熱しないでください。 - 記録室入口貯水池に外部参照電極を配置します。
注: 内部参照電極を用いた MEA チップが装備されていますが、これはカスタム記録室にカバーされます。したがって、外部参照電極を使用する必要があります。塩素処理を必要とせず使用する準備ができたので、飽和 KCl ペレットは最も実用的です。
12. MEA ライブ マッピング
- 一度 4AP アプライド レベルおよび記録温度が安定、必要ターンとして、血流と一時的にそれらを停止するオフの位置に吸引の活栓。
- 逆ガラス パスツール ピペットを用いた MEA 記録室に 1 つの脳のスライスをすばやく転送します。ファイアーポリッシュ カール パスツール ピペット (図 1) または柔らかいコンパクトな小さなブラシを使用して必要な MEA の記録領域の位置を調整します。脳スライスにホールド アンカー (図 1) を配置します。位置に戻って、活栓を回すことによって、血流と吸引を再起動します。
注:重要な!スライスを転送する必要がありますと 60 以内に、灌流を再起動 s や組織が死ぬことがあります。脳スライスが表面張力の違いによる脳スライスにアンカーを配置しながら移動するを防ぐために 4AP アプライドでスライス ホールド アンカーしておきます。MEA に脳スライスを確保するためアンカー ステンレス鋼ワイヤーとナイロン スレッド (図 1) を使用してカスタムすることができます。 または商業源から得られます。MEA チップを最終的な実験の設定はアンプのヘッドに接続図 1Eショー: カスタム アンカー記録室内 MEA チップで休んで脳スライスを押した。出口貯水池内吸引針 (青い矢印) の配置に対し、基準電極 (赤い円) と暖房カニューレ (赤矢印) をカバー チューブ PTFE は入口貯水池に配置されます。
- 逆ガラス パスツール ピペットを用いた MEA 記録室に 1 つの脳のスライスをすばやく転送します。ファイアーポリッシュ カール パスツール ピペット (図 1) または柔らかいコンパクトな小さなブラシを使用して必要な MEA の記録領域の位置を調整します。脳スライスにホールド アンカー (図 1) を配置します。位置に戻って、活栓を回すことによって、血流と吸引を再起動します。
- 倒立顕微鏡ステージ上に設置されたカメラを使用して脳スライスの写真を撮る。
- 電極をマップする GUI を開始するコンピュータ ・ ソフトウェアのスクリプトmapMEAを実行します。
注: カスタムメイドのソフトウェアは、脳スライスの特定の構造に対応する電極を選択するユーザーをことができます。この手順は、正しい経路をアクティブにし、電気刺激による発作活動を抑制することが重要です。

図 2: のための GUI のライブMEA 電極マッピング。(A) MEA に配置されている結合された海馬 EC スライスの模式図。このプロトコルで使用される既定の構造は、コルニュ ammonis 3 (CA3) enthorinal 皮質 (EC)、嗅周囲皮質 (PC) と subiculum (サブ)。参照電極は三角記号のマーカーとして描き出しました。点線で囲まれた電極は、イメージの矯正の XY 座標を表しています。(B) 画面は、GUI のライブの MEA マッピング中にキャプチャします。キャプチャの左のフロー チャートは、ステップバイ ステップの手順を示します。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
- 脳スライスの画像をロードする [参照] ボタンをクリックします。参照電極が (図 2 a、三角形マーク) MEA の左半分側の上の行に表示されることを確認します。ポインターをアクティブにするボタンをクリックし、イメージを矯正と電極マッピングの XY 座標をマークする配列の左端の行の上部と下部の電極を選択します。
- スライスの種類ドロップ ダウン メニューから選択、水平にします。既定の構造をチェックします。
注:新しい構造の入力ボタンを選択して、構造をカスタマイズすることが可能です。水平脳スライスの既定の構造は、図 2 aに描かれています。 - 脳スライスの写真の下の番号の押ボタンを使用して、投資収益率に対応する電極を選択し、割り当てる (図 2 b); 構造パネルの対応するボタンをクリックしてくださいそれぞれの投資収益率のためには、この手順を繰り返します。
- 保存ボタンを押す: ソフトウェアは、選択した電極や Roi レポート テーブルを含む#EXP_LabelledElectrodesをという名前の結果フォルダーを生成します。
13 てんかん活動記録と電気変調
- 録音の前に 5-10 分の記録チャンバー内を安定させるために脳のスライスを許可します。
- 自己校正と安定化させる刺激プロトコルの前に刺激の単位、少なくとも 10 分を入れます。刺激制御ソフトウェアを起動し、刺激および MEA アンプは正常に接続されている刺激制御ソフトウェアのメイン パネルの緑の LED によって示されることを確認します。特定の楽器や材料表を参照の追加の詳細については、ソフトウェアのマニュアルを参照してください。
- バイポーラで刺激を設定します。(Cf.セクション 12) スクリプトmapMEAにマップされているものの中で CA1/近位の subiculum (参考:24,25) の錐体細胞層との接触電極のペアを選択します。選択した電極の一つを刺激と同じ刺激装置チャネルの肯定的なプラグに他の電極の負のプラグに接続するのにワイヤを使用します。アンプのアース線を刺激の地面を接続別のワイヤを使用します。
- レコーディング ソフトウェアを起動します。データを取得するには、記録ソフトウェアのメイン パネルで再生ボタンを押します。レコードの少なくとも 4 発作放電。
注: 2 kHz のサンプリング周波数では、ハード ディスク領域の使用量を最小限に抑えながら公正な解決と電位を獲得ことができます。5 分記録ファイルは、〜 80 MB を取ります。高いサンプリング周波数が必要など、レコード刺激アイテムまたはマルチ ユニットの活動に。フィールド電位のみを観察するには、マルチ ユニットの活動を遮断する 300 Hz でライブのローパス フィルターを使用します。 - 刺激強度を決定します。
- 刺激コントロール ソフトウェアのメイン ・ パネルを使用して、正方形の相性正・負現在パルス持続時間 100 μ s/相をデザインします。
注:注意!直流刺激は、機器の損傷を避けるためにバランスの取れた充電パルスを必要とします。 - 最高の刺激強度を識別するために高速な入力/出力 (I/O) テストを実行します。5 のパルス間間隔を追加することによって手順 13.5.1 0.2 Hz 以下で設計されている刺激パルスを配信 s 以上刺激制御ソフトウェアの適切なフォームで。パルス振幅] タブで入力 100 μ A/相の初期パルス振幅と刺激は確実に海馬傍皮質の発作間欠期のようなイベントを呼び起こすことができるまでの各裁判で 50-100 μ A ステップを増やす (可視化された信号をチェック、レコーディング ソフトウェア) です。
- 刺激コントロール ソフトウェアのメイン ・ パネルを使用して、正方形の相性正・負現在パルス持続時間 100 μ s/相をデザインします。
- 大脳辺縁系発作原性の電気変調
- プログラムの利益の刺激のプロトコルを提供する刺激のユニット。I/O テスト中に識別される刺激の振幅を使用します。
注: 換起された応答の故障率は、引時間: 20% をする必要があります。 - 刺激を止めて後、は、(ステップ 13.4) のように、少なくとも 4 発作放電を記録することによって前刺激条件にネットワークの回復を確認します。
結果
6 × 10 のレイアウトと 500 μ m の電極間隔の平面測定が完全に脳スライス全体の記録領域にまたがるのでここで説明実験的プロトコルの理想的な記録デバイス (図 3 a、また17を参照してください)。穴あき Mea (pMEAs) は、組織の酸素化を改善することが望ましいだろうが、その記録領域が小さすぎます (~ 2 mm 径)。これは海馬と海馬傍皮質によって生成された電気信号の同時可視化計測はできません (データは表示されません;18を参照してください)。
何でこの体外モデルを使用して従来のフィールド電位観測され通常、アクティビティ7,の 3 種類で構成されて観測された 4AP 誘導てんかんパターン (図 3 a) を忠実に再現します。26: (私) 発作のようなのイベント (図 3 b矢印)、堅牢な長期的な (> 20 s, 範囲: 20-60 s) 強壮剤と間代コンポーネント (図 3); 発作活動のエレクトロ グラフの特徴に似ているイベント彼らは、海馬傍皮質内 3-5 分ごとに生成し、(図 3 D、矢印) 海馬歯状回、海馬形成を再入力(ii) 低速の発作間欠期のような放電 (図 3 b、EC トレース、黒いバー; で拡大図 3E) が短い (< 1 s) 海馬 EC 脳スライス標本の内で普遍的発作のイベントの間に生成される人口イベントゆっくりとしたペース (範囲 10-30 s) で発生して(iii) 高速発作間欠期のような放電 (図 3 b, CA3 トレース、黒いバー;図 3Eで拡大)、定期的な短い (< 1 s) CA3 海馬サブフィールド; によって生成される人口イベント高速 CA3 駆動間欠期イベントが海馬のループに沿って伝播し、抗 ictogenic 効果24 (示されていないデータ) を発揮シャファー側枝を保持する場合発作活動を防ぐために説明したプロトコルの目的のため、荒廃 CA3 出力が中断される (参考: 図 3 b、 E)。遅いペースで高速 CA3 駆動間欠期活動記録測定を使用してが発生することに注意する必要がある (〜 0.4 Hz) ガラス ピペットを使って記録従来フィールド ポテンシャルを用いた観察されたものよりも(~ 1 Hz の範囲: 0.5 - 2.0 Hz データは表示されません)。
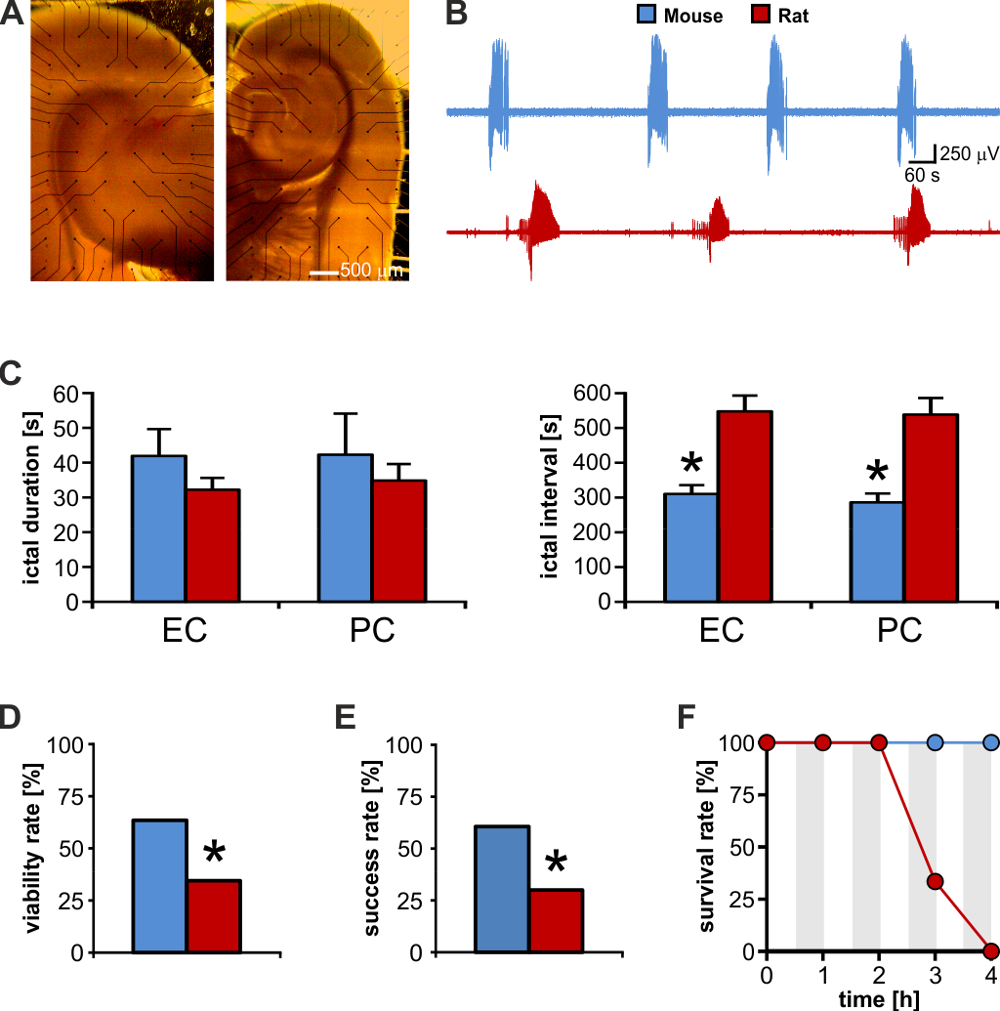
齧歯動物の脳スライスにおける電気ニューロモジュレーションの成功の結果は、準備24に含まれている地域の中で本質的な接続と共に特定の神経の細道27の活性化に依存します。我々 は雄 Sprague-dawley ラットから得られたスライスをテストして、CD1 マウスと我々 は、ラット脳スライスのサイズ、太さ、および組み込みの接続の間で最善のトレードオフを提供するのではなく、マウスを発見しました。まず、サイズが小さいおかげで彼ら合う市販 MEAs の小さな記録領域 (図 4 a、左: ラット、右: マウス);第二に、彼らは MEA の記録 (参考: 導入) に内在する、不利な条件に弾力の詳細: これらの 2 つの組織 (図 4 b) によって生成される発作のような放電が同様の期間の(図 4左)、その発生率はマウス脳切片で有意に短かった (n = 10 スライス種 2 対になっていない t-検定, p < 0.001;図 4右)。後者は、スピード アップ可能単一の実験的日中脳スライスの大きい数をテストする実験のプロトコル: 代表的な結果は、このセットの 2 種の脳スライスの同じような数をテストできる (ラット: n =58;マウス: n = 52)、マウスの数が (n = 18) ラット数倍未満 (n = 42)。
さらに、マウスの脳スライス高密度接続を提示する表示されます、したがってより持続的な電気ニューロモデュレーション (図 4) に対応する可能性が高い。したがって、この生体外モデルで発作原性の発作活動の制御を目的とした電気刺激のため生存率がラット脳組織よりもマウスのかなり大きい。この面は刺激実験ラット脳スライス対マウスを使用していくつかの持続的な刺激プロトコル(図 4 eが求めの有意に高い成功率にも反映)。実際には、マウス脳組織が良い 4 h (図 4 階) のテスト実験時間枠内で高い生存率が推奨する、持続的な電気刺激に耐えるように表示されます。従って、組み込み接続のより良い保全とともにサイズを小さく作るマウスの脳スライス MEA 海馬海馬傍ネットワーク相互作用の研究を目指した記録を実行し、評価する最高の候補者電気ニューロモデュレーション TLE に関連するプロトコル。
定期的なペーシング CA1/subiculum で配信制御、4AP 扱われる海馬 EC スライス準備24,25,27, 最も効果的な周波数27と 1 hz の辺縁系発作原性知られています。したがって、(例えば見なさい25) と他ニューロモデュレーション政策の効率性を評価するための肯定的な制御として便利ですも。MEA を (すなわち、外部刺激電極を必要とせず) から直接電気パルスが齧歯動物の脳組織の人口の応答を呼び起こすことができる前述のように、(また見なさい28)。脳スライス内の神経細胞ネットワークのための十分な保全、MEA と刺激電極対の適切な選択に脳スライスの正確な位置決めを追求できるように電気ニューロモデュレーション実験では,発作原性を効果的に制御します。図 5 aに示すように、正規の 1 Hz の周期記録と電気刺激の効果を全体で視覚化できる付加的な利点の刺激的なデバイスの両方平面測定を使用してプロトコルをペーシングを再現することが可能だ、従来のフィールド電位計測と比較して均等に間隔をあけられた地点数の増加と脳組織切片。図 5Bに示します n の結果の定量化 = 9 脳スライス、刺激の間の時間 (合計発作活動期間/観察時間25) を押収合計の大幅な削減を示す (1 つの方法-分散分析, F(df):6.84(2)、 p < 0.01、フィッシャーの LSD投稿アドホックテスト、保護された)。結果は、外部刺激電極24,25の文献と一致しています。
観測時間を押収合計が間隔で発作時のイベントに依存し、も刺激プロトコルの最小時間を決定を計算するには、自信を持って, 神経調節の効果を評価するため必要です。我々 は、少なくとも 4 発作放電 (およびしたがって少なくとも 3 発作イベント間隔) 記録が収集されたサンプルと必要なコレクション時間の間適切なトレードオフであることを見つけます。コントロールの状態 (すなわち、ない刺激) で観測時間は最初の測定の発作時イベントの発症と最後の 1 つの終了との間のラグです。ニューロモデュレーション, 中に観測時間は摂動はシステムで導入された時間の中で発生する現象を定量化する測定の目的は刺激プロトコル期間を等しくなります。コンピューティングと時間ではなく期間と発作時イベントの間隔を押収合計を比較することは、タイプ II のエラーを避けるために最も適切なパラメーターです。実際、発作活動の完全な抑制と共にも可能だいくつかのイベントではなく電気刺激中に発作時のイベントが生成されます 1 つだけ観測条件を制御されています。この場合、イベント間の間隔の測定ができないに対し刺激効果の推定量として発作の持続時間を測定を表さないニューロモデュレーション (すなわち, 発作活動の減少) の実際の結果です。
全体的にみて、結果はマウスの脳スライス MEAs の結合は、てんかん研究、てんかん治療のための DB のフィールドを事前に関連する信頼性の高いニューロモデュレーション研究を実行する貴重なツールであることを示します。さらに、ここで説明したプロトコル脳スライスの生存率が向上でき、異なる電気刺激の効果を比較するために、たとえば、必要がありますいくつかの時間 (図 6)、実験的セッションを追求パラダイム。

図 3: 平面の計測と可視化の典型的な 4AP 誘発てんかんパターン(A) マウスの脳スライス平面 6 x 10 MEA に配置されている (電極間距離: 500 μ m) と MEA の記録と視覚化 4AP 誘発てんかんパターンのサイド ・ バイ ・ サイドの概要。グリッド内の各升目脳スライス内の対応する位置で記録された活動に対応します。青い破線より明確に電極の行を参照してください。記録電極数は、それぞれの正方形の左上隅に青色で識別されます。灰色の正方形交差参照電極を表します。(B) 代表的なトレース セグメントより速い時間スケールで可視化した EC と CA3 のパターン。矢印は、発作時のイベントを示します。EC トレースでそれは低速の出現に感謝すること間歇放電 (黒いバー)、一方、CA3 サブフィールドは典型的な持続的な fastinterictal パターン (黒いバー) を生成します。(C) は、典型的な強直間代パターンを示す発作放電の記録を拡大しました。(D) (B) に赤いバーでマークされた EC (赤) と CA3 (黒) トレース セグメントに対応するプレ-ictal-に-発作時の遷移の高速スケール可視化。矢印は、発作時の放電の開始を示します。重ねられたトレースは、EC に由来する発作のイベントですが、その後 CA3 サブフィールドに伝達を強調表示します。(E) (B) の黒いバーでマーク発作間欠期の期間を拡大遅いの相関の欠如を強調すると fastinterictal パターンそれぞれ EC (赤) と CA3 (黒) によって生成されます。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 4: マウス脳切片ラット脳スライスよりも高い実験の出力を提供します。ネズミ (左) と一致した年齢のマウス (右) から (A) 脳をスライスします。ラット脳スライス、MEA 記録領域よりも大きいとその電気的活動が完全に可視化することはできません。(B) マウスとラットの enthorinal 皮質 (EC) によって生成された再発発作活動の直接的な視覚的比較。(C)マウスおよびラット脳スライス生成同様の期間の発作放電ですがこれらのイベント間の間隔は約 2 倍のラット脳組織。 * p < 0.05。(D) のラット、マウスに比較して発作原性を制御することを目的とした電気刺激実験のため実行可能な脳スライスの 2 倍より高い利回りを提供しています。Subiculum の電気刺激が人口 58 (34%) ラット脳スライス、52 (63%) の 33 ではなくマウスの脳スライスの唯一の 20 の海馬傍皮質の応答を呼び起こす (カイ2: 9.22;p = 0.002)。p < 0.05。(E) 実行可能なスライス間発作活動を制御することで成功率は 2 倍ラット脳スライス対マウス (マウス: 33 脳スライス、60% の 20: ラット: 20 脳スライス、30% の 6チー2: 4.67;p = 0.03)。p < 0.05。(F) マウス脳切片を耐えることができますには、持続的な電気刺激 (グレー網掛け領域) の長期の新紀元が繰り返されます。マウス脳組織がてんかんパターンの完全復旧を展示してラット脳組織の生存率は 3 h で劇的に落ちるに対し刺激撤退時にこの図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 5: 発作原性脳スライスを使用しての電気変調の代表的な実験は、高さと結合した4AP 誘発性発作活動 EC と PC の定期的な 1 hz (PP) をペーシング コントロール状態 (刺激) の間に (A) の録音 (刺激アーチファクトは切り捨てられます)、刺激撤退時に回復中。(B) 1 Hz で PP の合計時間の強制に及ぼす影響の定量化。p < 0.05。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 6: 4AP 誘発性発作活動の代表的な長期的な記録します。(A) トレース セグメント プロシージャをスライス脳後記録された 8 h と 4AP アプリケーションの次の 2 h。(B) 展開 (、) を長期観測時間未亡人を通して生成される発作活動の一貫性、手紙 b、およびcパネルによって識別される箱入り発作放電に対応するセグメントをトレースします。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
| ストック A | 化学 | 分子の重量 | 濃度 (mM) |
| 塩化ナトリウム | 58.44 | 1150 | |
| 溶剤:蒸留水 | KCl | 74.55 | 20 |
| 集中: 10 x | KH2PO4 | 136.1 | 12.5 |
| 貯蔵温度: 4 ° C | CaCl2* 2 H2O | 147.02 | 20 |
| 最大保存期間: 1 週 | D-グルコース | 180.2 | 250 |
| 注: 50 mm の膜フィルターを用いたフィルター | |||
| ストック B | 化学 | 分子の重量 | 濃度 (mM) |
| NaHCO3 | 84.01 | 260 | |
| 溶剤:蒸留水 | |||
| 集中: 10 x | |||
| 貯蔵温度: 4 ° C | |||
| 最大保存期間: 1 週 | |||
| 注: 50 mm の膜フィルターを用いたフィルター | |||
| 株 C | 化学 | 分子の重量 | 濃度 (mM) |
| KCl | 74.55 | 20 | |
| 溶剤:蒸留水 | KH2PO4 | 136.1 | 12.5 |
| 集中: 10 x | MgCl2* 6 H2O | 203.3 | 50 |
| 貯蔵温度: 4 ° C | MgSO4* 6 H2O | 246.47 | 20 |
| 記憶域の最大時間: 2 週 | CaCl2* 2 H2O | 147.02 | 5 |
| 注: 50 mm の膜フィルターを用いたフィルター | |||
| ストック M | 化学 | 分子の重量 | 濃度 (mM) |
| MgSO4* 6 H2O | 246.47 | 100 | |
| 溶剤:蒸留水 | |||
| 集中: 100 x | |||
| 貯蔵温度: 4 ° C | |||
| 記憶域の最大時間: 4 つの wks | |||
| 4AP ストック | 化学 | 分子の重量 | 濃度 (mM) |
| 4-アミノピリジン | 94.11 | 250 | |
| 溶剤:蒸留水 | |||
| 集中: 1000 x | |||
| 貯蔵温度: 4 ° C | |||
| 記憶域の最大時間: 2 週 | |||
| 注: 2-3 分の 30-60 分のなびくの渦 | |||
| 注: 注意!4 AP は有毒であり cinvulsant です。手袋を使用し、拡散や流出を避けるため。 | |||
表 1: 在庫ソリューション。
| アプライドを保持 | 記録アプライド | 切削アプライド | |||||
| 化学 | C [mM] | 化学 | C [mM] | 化学 | C [mM] | ||
| 塩化ナトリウム | 115 | 塩化ナトリウム | 115 | ショ糖 | 208 | ||
| KCl | 2 | KCl | 2 | KCl | 2 | ||
| KH2PO4 | 1.25 | KH2PO4 | 1.25 | KH2PO4 | 1.25 | ||
| MgSO4 | 1.3 | MgSO4 | 1 | MgCl2 | 5 | ||
| CaCl2 | 2 | CaCl2 | 2 | MgSO4 | 2 | ||
| D-グルコース | 25 | D-グルコース | 25 | CaCl2 | 0.5 | ||
| NaHCO3 | 26 | NaHCO3 | 26 | D-グルコース | 10 | ||
| L-アスコルビン酸 | 1 | L-アスコルビン酸 | 1 | NaHCO3 | 26 | ||
| L-アスコルビン酸 | 1 | ||||||
| pH | 7.4 | pH | 7.4 | ピルビン酸 | 3 | ||
| 浸透圧 | 300 mOsm/Kg | 浸透圧 | 300 mOsm/Kg | ||||
| pH | 7.4 | ||||||
| 浸透圧 | 300 mOsm/Kg | ||||||
表 2: ソリューションの構成。
| トラブルシューティング | |||||
| 問題 | 対策 | ||||
| 発作放電が 'チャンク' を見てください。 | 灌流率を減少させるおよび/または吸引針の位置を調整することによって脳スライス上アプライドの音量を下げます。 | ||||
| 発作活動なし | (1) CA3 駆動高速間欠期イベントは皮質に反映されませんを確認します。小さなメス刃 (n.10 など) を使用してまたは場合シャファー側枝を切断する針必要がありますが、スライス ホールド アンカーのナイロン スレッドを切らないように注意してください。ステレオまたは正立顕微鏡は、倒立顕微鏡は、このタスクを非常に困難に対し最適です。 | ||||
| スライスの生存を確認(2) : 強い電気的刺激が発作活動を引き起こすことができますか?4AP アプリケーションの約 2 時間まで待つ、発作時のアクティビティが発生しない場合、脳スライスを変更します。 | |||||
| 脳スライス ホールド アンカーを配置するときに移動します。 | (1)は優しくスライス アンカーを削除し、脳スライスの位置を変更します。 | ||||
| (2)は、スライス アンカーが 4AP アプライドの均等に濡れていることを確認します。 | |||||
| (3)は、mea 脳スライス接着を優先する録音室から 4AP アプライドを削除します。 | |||||
| (4)スライス ホールド アンカーの位置を変更します。 | |||||
| 転送された後 MEA チップを浮かぶ脳スライス | (1)には、パスツール ピペットで余分なアプライド優しく吸い出しなさい。 | ||||
| (2)の MEA チップは、再塗装する必要があります。 | |||||
| 電気刺激は、ネットワーク応答を発生しません | 刺激強度を増加、電極のペアを変更します。 | ||||
| 電気刺激は、人口近位しかしない遠位皮質の応答を引き出すことができます。 | 問題は貧しい接続または低すぎる刺激強度のための ikely です。いくつかの皮質のエリアのみでは、電気刺激を無効または有効に可能性があります。脳スライスを変更することをお勧めします。 | ||||
| 信号はノイズの多い | (1)は、参照電極を確認してください: 参照電極は、アプライドに完全に深くないことなど、ノイズの多いベースライン入口貯水池の不完全な塗りつぶしによって引き起こされることは時。 | ||||
| (2)は、機器の全体的な接地を確認します。 | |||||
| (3)一定、柔らかい吸引音を聞くために吸引針位置を調整します。吸引針を接地してください。 | |||||
| (4)は、綿棒を使用してエタノールで MEA 外部接点を清掃します。 | |||||
| (5)の MEA チップを破損する可能性があります: MEA チップを変更し、成果物と信号対雑音比をご確認ください。 | |||||
表 3: トラブルシューティングします。
ディスカッション
測定は、神経生理学の調査のための貴重なツールし、探査と開発の数十年のおかげで近年の成熟度に達しています。従来のフィールド電位計測と比較して測定は観測点と既知の電極間距離、神経ネットワークの相互作用を正確に特定することが重要である数の増加の大きな利点を提供しています。
MEA の記録技術は、パッチ ・ クランプ記録15、単一ニューロンと神経ネットワークの活動の関係を調査するなど、他の電気生理学のアプローチにも結合できます。さらに、電位とマルチ ユニットの活動を同時に可視化する可能性は小さい神経細胞集団の活動と集団的神経回路の相関に貴重な洞察力を提供できます。膜電位感受性色素、カルシウム イメージング、および光遺伝学29との組み合わせは、多面体のアプローチを用いた神経生理現象を推定できます。薬理学的研究に加えて記録と同じシステムでの刺激の両方の実行の可能性、MEA 記録技術非常に強力で汎用性の高い: たとえば、シナプス可塑性の現象を研究することが可能です、てんかん、各種脳疾患の治療に DBS に関連している記憶形成30、今回紹介したニューロモデュレーション プロトコルを使用してのコア。てんかん脳組織16に MEA 記録技術を正常に適用できる最近の証拠を示しててんかん研究、この破壊的な病気を根本的なメカニズムを理解する両方の非常に貴重な有用性それを改善する DBS アルゴリズムを微調整します。
ただし、MEAs 強制的に脳スライス、水中録音室の要件と脳をさせるための必要性のための '制限' 条件下での電気生理学実験の追求、固体基板上の残りの部分をスライス、微小電極統合されています。したがって、脳組織に録音の質を影響可能性があります十分な酸素の供給を受信できません。
ここで説明されているプロトコルは、信頼性の高い記録し、研究の急性の in vitro における 4AP モデルを用いた測定と結合した齧歯動物の大脳辺縁系ネットワークの発作原性と電気刺激の評価に関連する情報を提供する長期の新紀元を追求するにはDBS のポリシー。
従来のフィールド電位計測の使用に基づくてんかんの辺縁系ネットワークの電気ニューロモデュレーションに関する先行研究と同様の結果24,25; マウスまたはラット脳スライスを使用しています。しかし、ラットの脳組織の使用証明より挑戦的、合理的にマウス7から得られた脳組織に比べて弱い接続のため。MEA の技法に関してマウス脳切片は、サイズが小さいおかげで最適です。また、ラット脳組織対マウスの発作のような放電の発生の高い率を考えると、それはより速くより効率的なデータ収集は順番に、1 つの実験日全体に比べ脳スライスをテストしすることがでくことが可能使用される動物の数を減らします。
既存のメソッドの意義:
ここで説明されているカスタムのチャンバーを使用して記録された発作時の放電期間と従来の MEA リング商工会議所と同じ MEA 型 (平面微小電極、 cf. 17) を使用して観察された発生率に似ているように表示されます。しかし、それがため実験ニューロモジュレーションの成功を追求を妨げる可能性があります低灌流率 (1 mL/分)、と共に従来のラウンド記録室を用いたとき、脳スライス厚が大幅削減しなければならないと強調する必要貧しい人々 に接続します。
このプロトコルでは少量カスタム記録室パッチ ・ クランプ記録室のデザインに触発さは、MEA の録音の成功の追求のために不可欠な安定性と信頼性の層流を提供します。それはまた組織生存率と固有の接続との間の適正なトレードオフを達成するために 400 μ m まで脳スライス厚を増加させます。それは確かに認識されてチャンバ内録音ソリューションを層流が脳スライス生理望ましい温度、酸素の影響を受けませんので、循環型で観察される pH 勾配ラウンド記録部屋19,20,31 (のような市販の MEA で提供されている)。このようなグラデーションと脳スライスに有害も実験的先入観を紹介します。灌流中の適切な交換を可能にする高灌流率 (5-6 mL/分)16,31と共に比較的小さなボリューム (~1.5 mL) の記録室 (3 回/分)。カスタムの商工会議所は簡単にすることができます手頃な価格で商業ソースから取得または 3 D 印刷技術を利用して生成されます。他の研究は、低騒音の蠕動ポンプの助けを借りてから 400 μ m 厚脳スライス16高灌流率 (5-6 mL/分) を実施し、1 mL にアプライドのボリュームを保ちながら従来の MEA リング チャンバーを用いた MEA 録音を報告しています。しかし、著者が細胞外の K+濃度の有意な増加を含む ephaptic 機構の 4AP モデルよりも影響が少ない可能性がある発作原性,すなわち, 低 Mg2 +の別のモデルを使用しています。11,22. 高灌流率はアプライド16記録大幅に増加する必要があります蓄積された細胞外 K+のうち高速洗浄のための 4AP モデルでは望ましくないことがわかった。実際には、発作時のイベント登場より 'チャンク' ASCF 灌流の速度は 2-3 mL/分に増加したし、脳スライスはアプライド厚で水没した堅牢な強直間代コンポーネントを展示する本格的な放電に戻った時に復元するのに対し低灌流率 (1 mL/分) と薄くアプライド層組織表面のレベル (データは示されていない) に右の。このプロトコルで記述されているカスタム記録室灌流中に 3-5 回を交換が可能/1 mL/min の流速で分。したがって、脳スライスへの全体的な酸素供給がまだ安定した温度と高い信号対雑音比を記録を保証しながら比較的低灌流率でも強く向上しました。最も重要なは、生理学的な値に細胞外の K+濃度を維持することが可能です。
発作原性の 4AP モデル低下 Mg2 +または増加 K+などのアプライド イオン組成のどんな重要な修正は不要し、興奮性と抑制性の伝達をそのままに保つためのユニークな利点を提供しています12グルタミン酸と gaba 作動性てんかん同期 (cf. 7) ネットワークの両方の重要な役割に照らしててんかん研究で関連性の高いである側面。このプロトコルで使われる 4AP アプライドでは、生理的 K+濃度 (3.25 mM) と持株アプライド (1.3 mM と 1 mM) よりも若干低い Mg2 +濃度を含まれています。この濃度はまだ齧歯動物の脳脊髄液中の Mg2 +濃度の報告された生理学的な値の範囲内にあり、(例えば見なさい17,18) 多くの実験室で使用されます。説明プロトコルの目的に、我々 は 4AP の効果で支援するためにこのわずかな減少を好むことを発見しました。
前作には既に記録室17,18中置かれていたとき、脳スライスに 4AP が適用されています。実験者は 4AP アプリケーションとてんかんパターンの発症との間の時間遅延を観察する必要があります、しない限り、我々 はこのアプローチがかなり時間がかかるし、長期記録と刺激セッションを追求する適していないを見つけるこのプロトコルで説明します。32 ° C で 4AP で脳スライスの中古インキュベーションは、脳スライスは他の組織切片を用いた実験を追求しながらシリーズで前処理することができますので多く実験の時間を節約できます。
脳スライスの長期生存率は、長期的にネットワーク機能を分析すると便利かもしれません。また、反復的な刺激への回復力向上は、実験者は、統計の信頼性の同じ脳スライスで実行する必要がありますいくつかの刺激のプロトコルを比較したい場合の大きな利点のです。私たちの手で制御フェーズでそれぞれ先行 3 刺激プロトコルをテストする場合、回復期に続いて、実験は 3-5 h を最後可能性があります。このコンテキストでライブ MEA マッピングが正しい経路をアクティブにして抑制ではなく電気刺激による発作原性を支持するために重要です。ユーザーフレンドリーな GUI は、異なる脳構造の電極をマップする、簡単迅速かつ柔軟なツールを表します。商用ソフトウェアに反してプログラミングの基本的なスキルもカスタム MEA レイアウトを追加することが可能です。明るいフィールドに画像を得ることができます。従って、良好な解像度を持ち、顕微鏡に収まる任意の汎用カメラは適しています。これらは正立型顕微鏡を使用している場合は、組織の下に隠しだろうので脳スライスを基準電極位置を視覚化する倒立顕微鏡が欠かせません。ただし、ステレオ顕微鏡は、倒立型に加えては、実験者は、特定の神経経路を妨害するナイフ カットを実行する必要がある場合に推奨されます。
最後に、提案手法のさらなる利点は安価で比較的簡単なアセンブリのカスタム記録室と保持チャンバー、温かいお風呂とスライス アンカーとしての高価な不必要な使用のような必要なツールのほとんどの低騒音蠕動性ポンプ。
技術の制限:
MEA の記録は非常に遅い波、すなわち可視化を許可しません、DC 信号でシフトします。このようなベースラインのたわみは発作時の放電時間の漸近的に測定を助けるかもしれないし、最も重要な皮質拡散うつ病 (現象てんかん32の突然の予想外の死に関連して、共有されるを勉強する基礎です。間てんかんや片頭痛33)。
電気神経について脳スライスの生存率に影響を与えずにいくつかの刺激のセッションを実行できるようにここで説明されているプロトコル。20-45 分、以前作業25に報告したものと同様の最大 3 つの刺激セッションを正常に実行できます。脳スライス刺激セッションまたはより長い刺激プロトコル数の増加に耐える可能性がありますおそらくが脳スライスはこの点ではテストしませんでした。3 刺激プロトコルの数を制限して回避する長期大幅刺激撤退時に回復の不足までのこれらの制限条件下で維持される脳組織に重点を置くことができる (≥60 分) 刺激セッションをお勧めします。
プロトコルの中で重要なステップ:
いくつかの重要な要因には、レコーディングの成功を追求、MEA と結合した脳スライスにおけるてんかん活動の変調を妨げる可能性があります。ほかに使用される齧歯動物種と脳スライス、記録中に灌流率の品質、アプライドの流れのダイナミクス (すなわち層流円形と) 記録チャンバ内であることが回復の条件と。脳スライスの長期のメンテナンスだけでなく、4AP アプリケーションのモードは、最も重要な手順です。
浸漬保持チャンバーを用いた脳組織のネットワーク活動を維持し、4AP アプリケーションによる発作のような放電の誘導を支持するために室温で脳スライスを保存することが重要です。私たちの手で回復および 32 ° C でメンテナンス脳組織スライスの 3-4 時間以内に悪化してください。
脳のインキュベーションを室温で 4AP アプライドでスライスし、32 ° C での記録は、しかし、好ましくはないです。この状態で堅牢な再発発作放電を観察することで多くの難しさがわかりました。確かに、20-24 ° C の範囲で低温を弱めるかも 4AP 誘発発作原性、生体内および34 35を体外と両方を防ぐに報告されています。したがって、4AP インキュベーションおよび記録温度は 30-34 ° C に該当する必要があり、一致する必要があります。4AP と脳スライスの突然の暴露での興奮性亢進の同時誘導のためあまりにも多くのストレスの下で脳組織が室温から暖かい (32 ° C) 4AP アプライドへ置くことを避けるため、中間にあらかじめ温暖を実行する基本的なです。ステップと脳スライスは、このステップをスキップして 20-30 分の 4AP によって発作原性誘導に影響を与える可能性があります 32 ° C で持株アプライドで慣らします。
言及する必要がありますもう一つの側面は、スライス後、アプライド D グルコース濃度の重要性です。25 mM 濃度維持脳スライスよりも多くの実験室で使用されている 10 mM の濃度とは 2 倍速く発作活動の発生率も向上 (したがって、やすく実験の長いシリーズを追求するにはいくつかの脳スライス各日のプロトコル)。
最後に、スライスの処理中にいくつかの重要な細部は、言及する必要があります。まず、許可しない、そのままの脳と脳スライス切断アプライド寒さで凍結します。脳を低温 < 2 分は十分なマウス脳の小さいサイズを与えられました。切片は、室温にリンスのビーカーにバッファー トレイからすぐに移送すべき。切断を洗浄アプライドは高糖度が保持室のナイロン メッシュに固執するスライス脳になるそれ以外の場合非常に重要です。組織を効果的にすすぎにアプライドの持株を過度に汚染しないように転送ピペット内の切削アプライド コンテンツを最小限にすることが重要です。2 段階洗浄汚染を最小限に抑えることに役立ちます。スライスの手順が完了したら、組織切片に転送してくださいすぐにすすぎビーカーからソフト ナイロン メッシュのアプライドにより組織の酸素化、両側保持で中断された半分の方法、開催商工会議所。スライスの手順に従ってすべての段階に適用される同じ原理: 脳スライスはビーカーの底に座っているか、保持室の側面と接触することはできません、彼らは決してお互いに接触するはずもオーバー ラップ。
変更とトラブルシューティング:
アプライドの組成は、実験者のニーズに応じて変更すること。たとえば、特定の神経伝達物質や電気神経調節の効果イオン チャネルの貢献を分析する薬を追加可能性があります。さらに、ピルビンとアスコルビン酸 (参考: 表 2) 省略されるかもしれません、我々 を見つける強力な神経保護役割を出すが。表 3にこのプロトコルと、それらに対処する方法で発生することができます最も一般的な問題を報告する.
結論:
MEA の記録は間違いなく健康および病気の神経ネットワークの相互作用に対処するための貴重な技術であります。薬理学的研究に加えて、DBS てんかんと他の神経疾患に適用に関連する電気ニューロモデュレーション プロトコルを評価することが可能ですも。このプロトコルでは我々 は従来の細胞外フィールド電位計測で得られるものと同様の結果を持つ典型的な周期の刺激実験を再現することが可能、外部の刺激電極である示されています。業務用の使いやすい機器と高度なソフトウェア ツールの増加空室状況作る MEA 記録技術脳組織へとアドホックの刺激を提供するために、閉ループ刺激実験にも最適神経ネットワークの応答のフィードバック機構の貢献度を調査します。
開示事項
著者が明らかに何もありません。
謝辞
G. p. は欧州連合 MSCA-IF-2014 プロジェクト Re.B.Us、H2020 フレームワーク プログラムの下での総会 n.660689 によって資金を供給マリー キュリー スクウォドフスカやつです。GUI mapMEA は自由に ilaria.colombi@iit.it を作成するリクエストを承ります。
資料
| Name | Company | Catalog Number | Comments |
| Poly-D-Lysine | Sigma-Adrich | P7886 | Needed for MEA coating |
| NaCl | Sigma-Adrich | S9888 | Chemical |
| KCl | Sigma-Adrich | P9541 | Chemical |
| KH2PO4 | Sigma-Adrich | 795488 | Chemical |
| CaCl2*2 H2O | Sigma-Adrich | C3306 | Chemical |
| D-Glucose | Sigma-Adrich | RDD016 | Chemical |
| NaHCO3 | Sigma-Adrich | S5761 | Chemical |
| MgCl2*6 H2O | Sigma-Adrich | M2670 | Chemical |
| MgSO4*6 H2O | Sigma-Adrich | M5921 | Chemical |
| Sucrose | Sigma-Adrich | RDD023 | Chemical |
| Pyruvic Acid, 98% | Sigma-Adrich | 107360 | Chemical |
| 4-aminopyridine | Sigma-Adrich | A78403 | Convulsant drug |
| STG-2004 | Multichannel System | STG4004-1.6mA | 4-channel stimulus generator with voltage (±8 V) and current output (±1.6 mA) |
| MEA1060 | Multichannel System | N/A | MEA amplifier |
| planar MEA | Multichannel System | 60MEA500/30iR-Ti w/o ring | Must be without ring to allow using the custom recordingchamber |
| McRack | Multichannel System | Recording software | |
| McStimulus II | Multichannel System | STG4004 control software | |
| TC02 | Multichannel System | TC02 | 2-channel thermostat |
| PH01 | Multichannel System | PH01 | Heating perfusion canula |
| MPH | Multichannel System | MPH | Magnetic holder for PH01 and suction needle |
| Elastostil E43 | Wacker | E43 | Elastomeric sealant used to mount the custom recording chamber onto the MEA |
| MEA Custom Chamber | Crisel Instrument | SKE-chamber MEA | Custom recording chamber |
| Ag/AgCl electrode, pellet, 1.0 mm | Crisel Instrument | 64-1309 | Reference electrode for the custom recording chamber |
| Tergazyme | Sigma-Adrich | Z273287-1EA | Enzymatic cleaner |
| MATLAB | The Mathworks | Programming environment for electrodemapping | |
| VT1000S | Leica Biosystems | VT1000S | Vibratome |
| Warner Instruments | 64-1309 | Ag-AgCl Electrode Pellet 1.0 mm (E205). Reference electrode. |
参考文献
- Fisher, R. S., et al. Operational classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology. Epilepsia. 58 (4), 522-530 (2017).
- WHO. . Epilepsy Facts Sheet. , (2017).
- Fiest, K. M., Birbeck, G. L., Jacoby, A., Jette, N. Stigma in epilepsy. Curr Neurol Neurosci Rep. 14 (5), 444 (2014).
- Brodie, M. J., Barry, S. J., Bamagous, G. A., Norrie, J. D., Kwan, P. Patterns of treatment response in newly diagnosed epilepsy. Neurology. 78 (20), 1548-1554 (2012).
- Tanriverdi, T., Poulin, N., Olivier, A. Life 12 years after temporal lobe epilepsy surgery: a long-term, prospective clinical study. Seizure. 17 (4), 339-349 (2008).
- Dingledine, R. . Brain slices. , (1984).
- Avoli, M., et al. Network and pharmacological mechanisms leading to epileptiform synchronization in the limbic system in vitro. Prog Neurobiol. 68 (3), 167-207 (2002).
- Rutecki, P. A., Lebeda, F. J., Johnston, D. Epileptiform activity induced by changes in extracellular potassium in hippocampus. J Neurophysiol. 54 (5), 1363-1374 (1985).
- Mody, I., Lambert, J. D., Heinemann, U. Low extracellular magnesium induces epileptiform activity and spreading depression in rat hippocampal slices. J Neurophysiol. 57 (3), 869-888 (1987).
- Huberfeld, G., et al. Glutamatergic pre-ictal discharges emerge at the transition to seizure in human epilepsy. Nat Neurosci. 14 (5), 627-634 (2011).
- Pitkanen, A. . Models of seizures and epilepsy. , (2017).
- Rutecki, P. A., Lebeda, F. J., Johnston, D. 4-Aminopyridine produces epileptiform activity in hippocampus and enhances synaptic excitation and inhibition. J Neurophysiol. 57 (6), 1911-1924 (1987).
- Gross, G. W., Rieske, E., Kreutzberg, G. W., Meyer, A. A new fixed-array multi-microelectrode system designed for long-term monitoring of extracellular single unit neuronal activity in vitro. Neurosci Lett. 6 (2-3), 101-105 (1977).
- Pine, J. Recording action potentials from cultured neurons with extracellular microcircuit electrodes. J Neurosci Methods. 2 (1), 19-31 (1980).
- Reinartz, S., Biro, I., Gal, A., Giugliano, M., Marom, S. Synaptic dynamics contribute to long-term single neuron response fluctuations. Front Neural Circuits. 8, 71 (2014).
- Dossi, E., Blauwblomme, T., Nabbout, R., Huberfeld, G., Rouach, N. Multi-electrode array recordings of human epileptic postoperative cortical tissue. J Vis Exp. (92), (2014).
- Boido, D., et al. Cortico-hippocampal hyperexcitability in synapsin I/II/III knockout mice: age-dependency and response to the antiepileptic drug levetiracetam. Neuroscience. 171 (1), 268-283 (2010).
- Gonzalez-Sulser, A., et al. The 4-aminopyridine in vitro epilepsy model analyzed with a perforated multi-electrode array. Neuropharmacology. 60 (7-8), 1142-1153 (2011).
- Hajos, N., et al. Maintaining network activity in submerged hippocampal slices: importance of oxygen supply. Eur J Neurosci. 29 (2), 319-327 (2009).
- Ivanov, A., Zilberter, Y. Critical state of energy metabolism in brain slices: the principal role of oxygen delivery and energy substrates in shaping neuronal activity. Front Neuroenergetics. 3, 9 (2011).
- MultichannelSystems. . Manual PH01. , (2018).
- Zhang, M., et al. Propagation of epileptiform activity can be independent of synaptic transmission, gap junctions, or diffusion and is consistent with electrical field transmission. J Neurosci. 34 (4), 1409-1419 (2014).
- Panuccio, G., et al. In vitro ictogenesis and parahippocampal networks in a rodent model of temporal lobe epilepsy. Neurobiol Dis. 39 (3), 372-380 (2010).
- Barbarosie, M., Avoli, M. CA3-driven hippocampal-entorhinal loop controls rather than sustains in vitro limbic seizures. J Neurosci. 17 (23), 9308-9314 (1997).
- Panuccio, G., Guez, A., Vincent, R., Avoli, M., Pineau, J. Adaptive control of epileptiform excitability in an in vitro model of limbic seizures. Exp Neurol. 241, 179-183 (2013).
- Benini, R., Avoli, M. Rat subicular networks gate hippocampal output activity in an in vitro model of limbic seizures. J Physiol. 566 (Pt 3), 885-900 (2005).
- D'Arcangelo, G., Panuccio, G., Tancredi, V., Avoli, M. Repetitive low-frequency stimulation reduces epileptiform synchronization in limbic neuronal networks. Neurobiol Dis. 19 (1-2), 119-128 (2005).
- Steidl, E. M., Neveu, E., Bertrand, D., Buisson, B. The adult rat hippocampal slice revisited with multi-electrode arrays. Brain Res. 1096 (1), 70-84 (2006).
- Pulizzi, R., et al. Brief wide-field photostimuli evoke and modulate oscillatory reverberating activity in cortical networks. Sci Rep. 6, 24701 (2016).
- Liu, M. G., Chen, X. F., He, T., Li, Z., Chen, J. Use of multi-electrode array recordings in studies of network synaptic plasticity in both time and space. Neuroscience Bulletin. 28 (4), 409-422 (2012).
- Maier, N., Morris, G., Johenning, F. W., Schmitz, D. An approach for reliably investigating hippocampal sharp wave-ripples in vitro. PLoS One. 4 (9), e6925 (2009).
- Aiba, I., Noebels, J. L. Spreading depolarization in the brainstem mediates sudden cardiorespiratory arrest in mouse SUDEP models. Sci Transl Med. 7 (282), 282ra246 (2015).
- MA, R., Avoli, M., Noebels, J. L., Rogawski, M. A. . Jasper's Basic Mechanisms of the Epilepsies . , (2012).
- Motamedi, G. K., et al. Termination of epileptiform activity by cooling in rat hippocampal slice epilepsy models. Epilepsy Res. 70 (2-3), 200-210 (2006).
- Yang, X. F., Duffy, D. W., Morley, R. E., Rothman, S. M. Neocortical seizure termination by focal cooling: temperature dependence and automated seizure detection. Epilepsia. 43 (3), 240-245 (2002).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved