
Method Article
פיצול-BioID — ניתוח פרוטיאומיה מבנית של חלבון ספציפי ההקשר מתחמי הסלולר בסביבתם הטבעית
In This Article
Summary
אנו מספקים פרוטוקול צעד אחר צעד עבור פיצול-BioID, שיטת שברי-קומפלמנטציה מבוסס על טכניקת קרבה-תיוג BioID. מופעל על האינטראקציה של שני חלבונים נתונה, היא מאפשרת ניתוח פרוטאומיקס של חלבונים תלויי-ההקשר מתחמי הסלולר בסביבתם הטבעית. השיטה היא פשוטה, חסכונית ודורש רק ציוד מעבדה סטנדרטיים.
Abstract
להשלמת הזיקה הקיימת טיהור (AP) מתקרב לשם הזיהוי של אינטראקציות חלבון-חלבון (PPI), אנזימים הוכנסו המאפשרות תיוג תלויי-הקרבה של חלבונים בתאים חיים. אחד כזה האנזים, בירה * (בשימוש הגישה BioID), המעביר את biotinylation של חלבונים בטווח של 10 ננומטר. לפיכך, כאשר דבוקה חלבון עניין הביע בתאים, היא מאפשרת תיוג של proximal חלבונים בסביבתם הטבעית. בניגוד AP המסתמך על הטיהור של מתחמי חלבונים מורכבים, BioID מזהה חלבונים שסומנו בתוך התאים, לא משנה אם הם עדיין אינטראקציה עם החלבון עניין כאשר הם מבודדים. מאז זה biotinylates הפרוקסימלית חלבונים, אחד יכולים יתר על כן לנצל זיקתו יוצאת דופן של streptavidin עבור ביוטין לבודד אותם ביעילות רבה. בעוד BioID מבצע יותר טוב AP מזהים ארעי או אינטראקציות חלשות, שתי הגישות ספקטרומטר AP - ו BioID-המונית מספקים סקירה של כל האינטראקציות האפשריות שיש חלבון נתון. עם זאת, הם אינם מספקים מידע על ההקשר של כל PPI שזוהה. ואכן, רוב החלבונים הם בדרך כלל חלק מספר מתחמי, המייצגים שלבים ברורים ההבשלה או יחידות פונקציונליות שונות. כדי לטפל מגבלה זו משותפת של שתי השיטות, שתכננו assay קומפלמנטציה של חלבון-שברי בהתבסס על האנזים בירה *. ב הזה assay, שני קטעים לא פעיל של בירה * ניתן להרכיב מחדש לתוך אנזים פעיל כאשר הביא בסמיכות שני חלבונים שמעצבת שאליו הם מקובעים. וזמינותו פיצול-BioID וכתוצאה מכך ובכך מאפשר תיוג של חלבונים להרכיב סביב זוג של חלבונים אינטראקציה. בתנאי שני אלה רק אינטראקציה בהקשר נתון, פיצול-BioID מכן מאפשרת הניתוח של יחידות פונקציונליות ספציפית תלויי-ההקשר הסלולר בסביבתם הטבעית. כאן, אנו מספקים פרוטוקול שלב אחר שלב ולבחון חל פיצול-BioID זוג של חלבונים אינטראקציה.
Introduction
כפי ביותר תאיים מבוצעות על ידי חלבונים באופן דינאמי קומפלקסים macromolecular, הזיהוי של אינטראקציות חלבון (PPI) היא מאמץ גדול במחקר ביו-רפואי. אכן, PPI הם לעתים קרובות deregulated במחלה, מייצגים מטרות אפשריות עבור הרפוי1. הנפוצים ביותר שיטת הזיהוי של PPI הוא הגישה טיהור (AP) זיקה שבו, בעקבות פירוק תאים, חלבון עניין מטוהר במיוחד על מטריצה, חלבונים הקשורים לאחר מכן מזוהים באמצעות ספקטרומטר מסה (MS). בעוד AP-MS זה בגישה רב-עוצמה, זה בדרך כלל אינו מבצע גם על מתחמי מסיסות חלבון, האינטראקציות מאוד ארעי או PPI הדורשים מבנה subcellular ללא פגע. יתר על כן, הפרשנות של הנתונים יכול להיות מסובך באופי הדינמי של רשתות PPI, כמו חלבון יחיד היא לעתים קרובות חלק מספר מתחמי חלבון ברורים.
תיוג-קרבה טכניקות כגון BioID2 או3,APEX24 פותחו לאחרונה כדי להתמודד עם המגבלות של הגישות AP-MS. ב- BioID, האנזים בירה * (תואם ל- variant G115R של האנזים e. coli פראי סוג) מזרז היווצרות יציב biotinyl-AMP (ביו-AMP) זה יכול להגיב עם ראשי אמינים. בניגוד האנזים פראי סוג, זה שומר על ביו-AMP במרכזו פעיל, בירה * משחררת ביו-AMP ומאפשר שלה דיפוזיה לסביבת השכנה שלה. לפיכך, כאשר דבוקה חלבון עניין והביע בתאים, חלבונים צינתור יכול להיות biotinylated בתוך טווח משוער של 10 nm5. אלה מסומנים proximal חלבונים הם מבודדים על ידי הנפתח streptavidin ואז המזוהה על-ידי MS. בניגוד AP-MS, BioID דורש את הביטוי של חלבון פיוז'ן. זה ובכך ניתן להחיל רק חלבונים שתפקידם היא לא הקשו על-ידי תיוג. יתר על כן, מהירות תיוג היא איטית, בדרך כלל 6-24 h2,6, ביצוע זיהוי חלבונים קצרת ימים מאתגרים. ובכל זאת, בהשוואה ל- AP-MS, BioID-MS מציע מספר יתרונות מרכזיים: הראשון, שלה לוכדת האינטראקציות בסביבה התאית הטבעית שלהם; שנית, שכותרתו חלבונים מאשר מתחמי שהורכב מבודדים בעקבות פירוק התא; שלישית, streptavidin pulldowns מאפשרות שימוש denaturing מאגרי ותנאים קשים כביסה. לכן, השיטה היא יותר רגיש כדי לזהות ארעי או אינטראקציות חלשות7 או האינטראקציות המתרחשות על ספציפי, קשה לבודד את מבנה subcellular8.
עם זאת, רוב החלבונים בדרך כלל מהווים חלק מתחמים גדולים יותר ניתן לשפץ על פי רמזים הסלולר או לפונקציה צריך להתבצע. ומכאן, חלבון יחיד הוא בדרך כלל חלק מספר מתחמי, המייצגים יחידות פונקציונליות נפרדים, הקשורים ברורים ו/או חופפים PPI. שתי הגישות לתת סקירה כללית של כל שיוכי שחלבון נתון ייתכן, אך הם נכשלים להתייחס בהקשר של הפרט PPI. כדי להגדיל את הרזולוציה של האחרון, עיצבנו של חלבון-שברי קומפלמנטציה assay (PCA) יכולים אילו שני קטעים לא פעיל של בירה * (NBirA *, המכיל את התחום קטליטי, ו CBirA * שניתן להציג בתור התחום הפעלה מחדש.) לכנס שוב לתוך אנזים פעיל כאשר הביא בסמיכות על ידי שניים אינטראקציה חלבונים9. וזמינותו פיצול-BioID וכתוצאה מכך מתמקד biotinylation תלויי-הקרבה של חלבונים להרכיב סביב זוג שמעצבת חלבונים, ובכך מאפשרת זיהוי הקשר מכלולים התלויים חלבון. לאחרונה להדגים את הכוח רזולוציה יוצאת דופן של פיצול-BioID על-ידי פענוח כשני מתחמים נפרדים חלבון מעורב בתיווך miRNA ג'ין להחרשת מסלול9.
בסך הכל, assay יחיד ופשוט, פיצול-BioID מאפשרת גילוי והקצאת במיוחד PPI ליחידות פונקציונלי מוגדר שבו חלבון נתון הוא מעורב, בתנאי שחלבון אינטראקציה נוספת של החלבון המתאים מורכבים ידוע.
Protocol
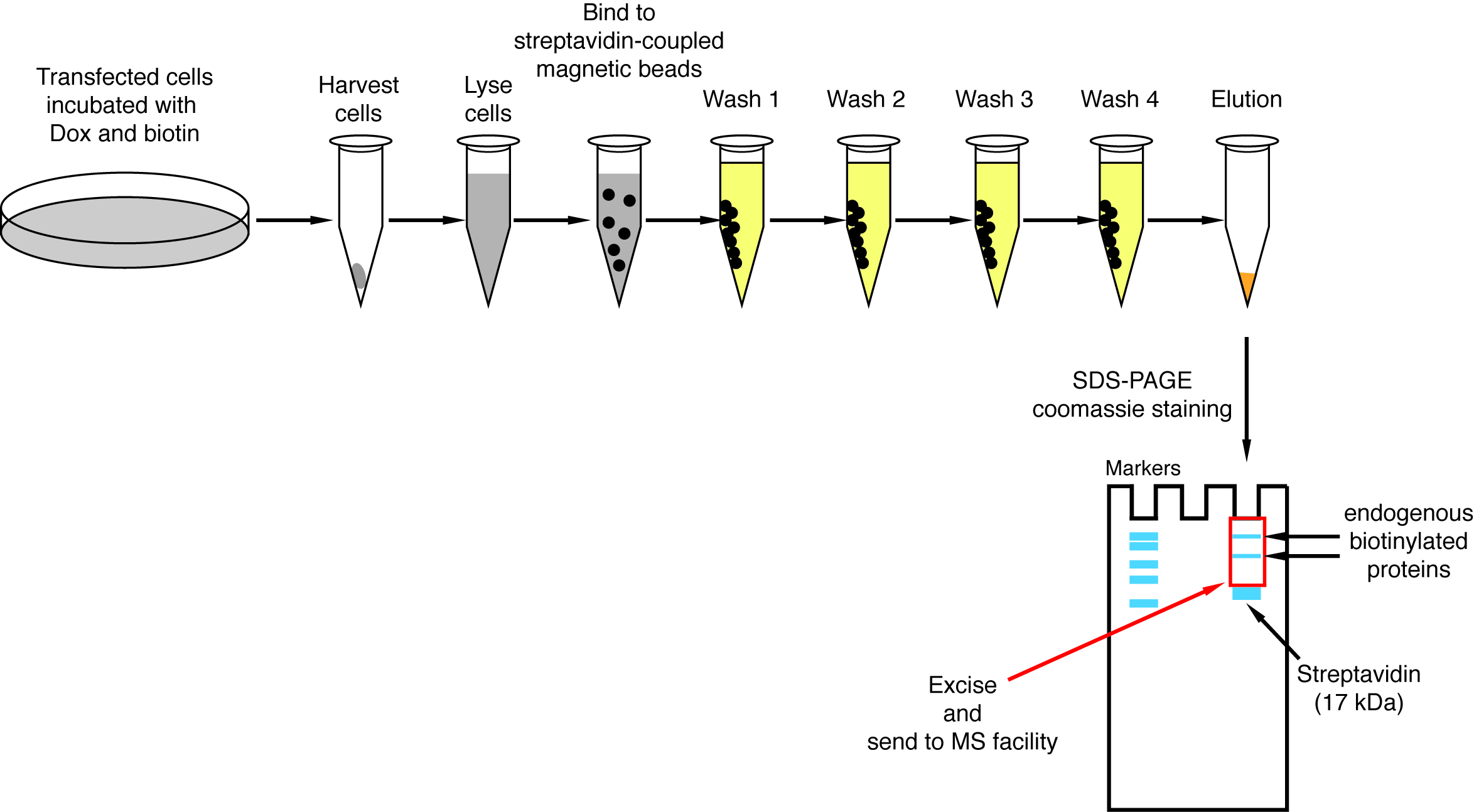
הערה: סקירה כללית של השיטה מוצג על איור 1.
1. תכנון של האסטרטגיה שיבוט
- בחר שני חלבונים putatively שמעצבת להיבדק.
הערה: כל אחד שני החלבונים נכרכים למקטע אחד של פיצול-BioID: NBirA * או CBirA *. כפקד שלילי, פיוז'ן החלבונים CBirA * ייבדק עם NBirA * דבוקה של חלבון פלואורסצנטי ירוק (NBirA *-GFP). בתור פקד נוספים, NBirA * fusions עשוי להיבדק לבד, בלי cognate CBirA * היתוך או בשילוב עם CBirA * דבוקה של חלבון לא קשורים. מומלץ לא להשתמש CBirA *-GFP כפקד שלילי כפי שהיא הוצגה להוביל באופן עקבי על רקע העיקריים בשילוב כל NBirA * פיוז'ן חלבון9. ייתכן שהגורם של התבוננות זו רמת הביטוי של CBirA *-GFP, גבוה בהרבה מאשר כל CBirA * פיוז'ן חלבונים אחרים השתמשנו עד כה, שעשוי להוביל reassociation משמעותית עם השבר NBirA *. -
ברגע נבחרו שני חלבונים, בדוק את הספרות כדי למצוא אם שנינו כבר בהצלחה תויגו במחקרים פונקציונלי (למשל כמו חלבון כימרי מתויג GFP).
- אם קיימים מחקרים כאלה, לציין את המיקום של התג (-או C-אמיני) ולהשתמש באותו הכיוון כדי לתייג את החלבונים של עניין עם שרידים פיצול-BioID.
- אם אף מחקר כזה קיים, בונה תוכנית קידוד עבור שני חלבונים מתויג ב ו C-אמיני ולתכנן assay כדי לבדוק את הפונקציונליות של החלבונים פיוז'ן (לדוגמה, ניסוי הצלה בקו הסלולרי מרוקן זמן החלבון WT).
הערה: בעת זיווג שני חלבונים היתוך באמצעות פלסמידים המתואר באיור 2 אשר קידוד linkers גמיש ארוך, הכיוון של החלבונים פיוז'ן (בירה * שברי מאוחה במעלה או במורד הזרם של החלבונים עניין) בדרך כלל לא משנה . ואכן באמצעות FRB ו FKBP בתור מודל חלבונים, זה הוצג כי כל ארבע איטרציות אפשרי (שני קטעים ב- N-טרמיני, הן ברמה C-טרמיני, אחד בכל קצה אמיני והשני קצה קרבוקסילי, ולהיפך) תשואות דומות biotinylation9.
- עיצוב תחל כדי להגביר את ORFs של שני חלבונים עניין לקראת שיבוט לתוך פלסמידים פיצול-BioID. שים לב כי המסגרות תרגום זהים השברים בירה *. אם משתמש של פלסמידים המתואר באיור2, להשתמש את אנזימי הגבלה PmeI, פאצי לבנות CBirA * הפיוז'ן, אנזימים הוא ו- MluI עבור NBirA * הפיוז'ן.
הערה: פלסמידים מתואר באיור 2 נושאים אלמנט מגיב ט דו כיווני ולצדו F3/והשפלתם רקומבינציה אתרים. פעולה זו מאפשרת את הביטוי שיתוף מוסדר בערך באותה רמה של שני חלבונים פיוז'ן של פלסמיד אותו10, ועל האפשרות לבנות במהירות שורות תאים יציב על-ידי המרת קלטות בתיווך רקומבינציה (RMCE). פלסמידים אלה, NBirA * כולל תג myc בזמן CBirA * יש תג דגל. ORFs ניתן כמובן גם להיות משובטת על פלסמידים בודדים עם היזמים מכוננת.
שלב קריטי: בעת בדיקת שני חלבונים אינטראקציה, מומלץ להשוות NBirA * דבוקה חלבון 1 בשילוב עם CBirA * דבוקה חלבון 2, NBirA * דבוקה חלבון 2 בשילוב עם CBirA * דבוקה חלבון 1. ואכן, הוא לעתים קרובות ציין כי איטרציה עובד טוב יותר מאשר השני.
2. שיבוט את ORFs של הגנים של עניין לתוך פלסמיד פיצול-BioID
הערה: בדוגמה זו, שני חלבונים יכולים להיות מתויגים על קצה אמיני נחשבים. ארבעה תנאים נבדק, לעומת תאים שאינם transfected (טבלה 1).
- השתמש תגובת שרשרת פולימראזית (PCR) כדי להגביר את ORFs של שני החלבונים להיבדק עם תחל המתאים. עיצוב תחל להציג האתרים סייג הוא, MluI של NBirA * פיוז'ן החלבונים, ואת האתרים פאצי, PmeI של CBirA * פיוז'ן החלבונים.
הערה: ה-PCR יכול להתבצע עם מסחריות, מעדיפים הגהה, ה-DNA פולימראז. בצע את הפרוטוקול הסטנדרטי של בהוראות ולהתאים אותו טמפרטורת ההיתוך צבעי בסיס ואורך ORF הגברה על פי הנחיות היצרן. -
Subclone של ORF הראשון
- לעכל פלסמיד (μg על 2) והן מוגבר-PCR ORF1 (כדי להיות דבוקה NBirA *) ועם הוא MluI. לבצע תגובות מערכת העיכול באמצעות μL 1 של כל אנזים, מעורבב עם נפח מתאים של מאגר ה-DNA ותגובה של הנפח הכולל של 20 μL צינור 1.5 מ"ל לשעה ב 37 º C.
- להפעיל שתי דגימות עיכול על 1% agarose-טה DNA ג'ל. בלו הלהקות המתאים פלסמיד מעוכל, ORF1 עם האזמל נקי, העברת צינורות 1.5 mL.
- לטהר שתיהן באמצעות ערכת חילוץ DNA סטנדרטי.
- מאתרים ומפסיקים פלסמיד מעוכל, ORF1 בעזרת ריאגנטים סטנדרטי.
- אם באמצעות ערכת מצדו המצוין בטבלה של חומרים, מאתרים ומפסיקים 100 ננוגרם של ה-DNA המכיל חמש זמן עודף טוחנת להוסיף על פלסמיד ב הנפח הכולל של 4.5 μL. מוסיפים 5 μL 2 x T4 ליגאז מאגר 0.5 μL של T4 DNA ליגאז הערכה מצדו. ביצוע התגובה מצדו צינור 1.5 מ ל 10 דקות בטמפרטורת החדר (RT).
- להפוך DH-5α e. coli המוסמכת תאים סטנדרטיים (המוכנים לפי שיטה של ינו11).
- ΜL מיקס 3 התגובה מצדו עם 50 μL של תאים המוסמכים mL 1.5 צינור על קרח, תקופת דגירה של 30 דקות העברה לתאים חום לחסום מוגדר כ- 42 ° C עבור s 30-45, ואז דגירה על קרח 2 דק להוסיף 250 μL של טרום ומחוממת (37 ° C) lysogeny מרק (LB) בינוני וצלחת 100 μL של התאים על צלחת מראש ומחוממת (37 מעלות צלזיוס) המכיל אמפיצילין LB-אגר. דגירה התאים בן לילה ב 37 º C.
- ביום הבא, לבחור ארבע עד שש מושבות דגירה אותם ב 37 מעלות צלזיוס, 180 סל ד, לינה 3 מ ל LB בינוני המכיל אמפיצילין-1 μg⋅mL 100 צינור 15 מ"ל.
- לבודד את פלסמידים מן המושבות שנאספו באמצעות ערכת ה-DNA MiniPrep רגיל.
- לוודא את נכונות פלסמידים מאת סנגר רצף באמצעות פריימר הפוך את קלטת 2 (טבלה 2).
- ברגע פלסמיד המכיל את ORF הראשונה התגלתה, subclone של ORF השני בתוך פלסמיד את ביצוע הפעולות כשלב 2.2 אבל באמצעות אנזימים PmeI ו פאצי.
- רצף של פלסמידים הנוצרת באמצעות פריימר הפוך את קלטת 1 (טבלה 2).
3. בדיקה של החלבונים פיוז'ן
הערה: ההוראות הבאות הן של פלסמידים ביטוי inducible כפול (איור 2), הלה-11ht תאים, קו תא הלה-CCL2 subclonal, stably לבטא את מפעיל שעתוק במהופך שבשליטת טטרציקלין rtTA-M2, המכיל לוקוס RMCE12. מדיום הגידול עבור תאים אלה הוא של Dulbecco ששינה נשר בינוני (DMEM) המכיל 10% ללא טטרציקלין עוברית סרום שור (FBS). בעת שימוש בסוג אחר של התא, מדיום הגידול ותנאים זריעה מדויקת יצטרכו להתאים.
-
תרביות תאים ארעית
- תאי זרע-ריכוז של עונה 1 פרק 105 תאים ב- 2 מ לכל טוב של צלחת 6-ובכן יום לפני תרביות תאים, דגירה התאים בן לילה ב 37 ° C, 5% CO2 בחממה התרבות תאים.
- ביום של תרביות תאים, להסיר את המדיום של כל טוב והחלף 2 מיליליטר בינוני טריים.
- הכינו את התגובות ארבעה תקנים על פי טבלה 1. עבור כל טוב transfect, להוסיף 6 µg של polyethylenimine µg 3 של פלסמיד DNA צינור סטרילי 1.5 mL ולאחר מילוי כדי µL 500 DMEM בינונית ללא סרום.
- דגירה כל תערובת תקנים לפחות 5 דקות בטמפרטורת החדר לפני הוספת טיפה חכם מכל קידוח. דגירה התאים בן לילה ב 37 מעלות צלזיוס, 5% CO2.
-
אינדוקציה ולקרוא קרבה
- יום אחרי תרביות תאים, להסיר את המדיום והחלף בינוני בתוספת ביוטין-50 מיקרומטר לעורר biotinylation ו דוקסיציקלין-200−ng⋅mL1 כדי לעודד את הביטוי של החלבונים פיוז'ן. דגירה את התאים עבור פחות 20 h ב- 37 מעלות צלזיוס, 5% CO2.
- כדי להפוך את פתרון מניות של ביוטין, להמיס ביוטין-50 mg⋅mL-1 (שיתאימו ca. 200 מ מ) ב- 2 מ' אמוניה מימית. ברגע זה היא התפרקה לחלוטין, למהול אותו ל-50 מ מ ב- 500 מ מ Hepes, pH 7.4, ולאחר מכן להתאים את ה-pH ל 7.4 עם HCl. Aliquot ולאחסן 1,000 וכתוצאה מכך x פתרון מניות ב-20 ° C. להמיס דוקסיציקלין mg⋅mL 10-1 70% אתנול, בחנות microtube קאפ בורג ב-20 ° C בחושך.
- יום אחרי תרביות תאים, להסיר את המדיום והחלף בינוני בתוספת ביוטין-50 מיקרומטר לעורר biotinylation ו דוקסיציקלין-200−ng⋅mL1 כדי לעודד את הביטוי של החלבונים פיוז'ן. דגירה את התאים עבור פחות 20 h ב- 37 מעלות צלזיוס, 5% CO2.
-
הכנה lysate תא
- לשטוף את התאים פעם אחת עם 1 מ"ל של קור (4 ° C) באגירה פוספט תמיסת מלח (PBS).
- כדי כל טוב, להוסיף 100 µL מאגר פירוק (50 מ מ טריס pH 7.4, 150 מ מ NaCl, 2 מ מ EDTA, 0.5% NP-40, 0.5 מ מ DTT, מעכבי פרוטאז).
- לקצור את התאים עם תא להרביץ, העברת צינורות 1.5 mL.
- Centrifuge את הדגימות ב x 14,000 g 10 דקות ב 4 ° C כדי להסיר שאריות תאים.
- להעביר את supernatants צינורות טריים ומניחים על קרח.
- לקבוע את כמויות חלבון עם וזמינותו ברדפורד.
-
מרחביות-לזיהוי בג'ל (עמוד), סופג המערבי
- הכנת ג'ל מרחביות-לזיהוי.
- עבור כל פינה lysate, היכונו מדגם דף של חלבון 30 µg (מינימום 15 µg) סך של µL 30 מאגר העמסה מרחביות. ואז לטעון כמויות שוות של כל דגימה (20 µL טוב) על הג'ל מרחביות-לזיהוי והמשך אלקטרופורזה.
- לאחר אלקטרופורזה, להעביר את החלבונים fractionated קרום חשופה PVDF פלורסצנטיות נמוך באמצעות כל פרוטוקול סטנדרטי.
הערה: עבור ניתוח חלבון biotinylation, זמן העברת 10 דקות עם תוכנית "MW גבוהה" מהיר להעביר חצי יבש המערבי סופג את המכשיר בדרך כלל עובד היטב. - לאחר המעבר, לחסום את הקרום ב- 5% חלב יבש ב- PBS למשך 30 דקות ב- RT.
- דגירה קרום למשך 30-60 דקות ב RT עם פלורסנט streptavidin 1:15,000 מדולל תרכיב ב- PBS המכיל BSA 2% ו- 0.1% Tween-20.
- לשטוף את הקרום שלוש פעמים, כל 10 דקות, עם PBS המכיל 0.1% Tween-20, ולאחר מכן יותר עם PBS.
- סרוק את הקרום על מערכת הדמיה של הסורק זריחה.
הערה: חור טיפוסי תספיג מוצג באיור3. ההליך הנ ל מתאר של זיהוי מבוסס פלורסנט אך זיהוי מבוסס-הפריה חוץ גופית באמצעות streptavidin מצמידים HRP עובד לא פחות טוב.
4. פיצול-BioID ללימודי פרוטאומיקס
הערה ביקורתית: לניתוח המוני spectrometric הסופית, כל השלבים הבאים הם להתבצע בתנאים ללא קרטין, כל חומר, ריאגנטים צריך להיות כמו קרטין-חופשית ככל האפשר.
- תרביות תאים ארעית
- יום לפני תרביות תאים, זרע שלושה-ארבעה ס מ 10 לוחות עבור כל תנאי-ריכוז של תאים5 8 x 10 מ"ל לכל צלחת, דגירה התאים בן לילה ב 37 ° C, 5% CO2 בחממה התרבות תאים.
- ביום הבא, להכין תערובת בסיס תקנים עבור כל תנאי תרביות תאים: עבור לוחות שלוש לכל תנאי, 36 µg של polyethylenimine, µg 18 של פלסמיד DNA מומס 900 µL ללא סרום DMEM. דגירה בכל מיקס מאסטר לפחות 5 דקות ב- RT.
- בינתיים, להחליף את המדיום של כל מנה בינונית טרייה, ולאחר מכן להוסיף 300 µL הירידה תערובות תרביות תאים חכם כל צלחת.
- דגירה התאים בן לילה ב 37 מעלות צלזיוס, 5% CO2.
- אינדוקציה ולקרוא קרבה
- יום אחרי תרביות תאים, להעביר את התאים מנות 15 ס מ. עבור כל מנה, להסיר את המדיום, לשטוף את התאים עם 7 מ של PBS, להוסיף 1.5 מ של פתרון טריפסין-EDTA, תקופת דגירה של 5 דקות ב- RT. להוסיף 3.5 מ"ל של מדיום הגידול resuspend התאים ולהעביר התליה תא ל צלחת 15 ס מ מלא עם 20 מ של מדיום הגידול בתוספת ביוטין-50 מיקרומטר לעורר biotinylation ו דוקסיציקלין ב 200 ng⋅mL− 1 (ריכוזים הסופי) כדי לעודד את הביטוי של החלבונים פיוז'ן.
- דגירה את התאים עבור פחות 20 h ב- 37 מעלות צלזיוס, 5% CO2.
- איסוף ואחסון של תאים
- לשטוף את התאים פעמיים עם PBS, ולאחר מכן להוסיף 1.5 מ של PBS כל צלחת ולקצור את התאים עם דבקנית.
- להעביר את התאים שנקטפו התואם תנאי אחד לרכבת התחתית 15 מ"ל, מסיק מהם על ידי צנטריפוגה x 1200 גרם, 5 דקות, 4 מעלות צלזיוס.
- להסיר את supernatants, הצמד להקפיא את כדורי חנקן נוזלי, אז החנות ב 80 ° C עד עיבוד נוסף.
הערה: לחלופין, תאים יכולים גם להיות מנותק על-ידי trypsinization, שנבצרו מדיום הגידול, הועבר צינור 15 מ"ל ושטף בפעם השלישית עם PBS לפני ההקפאה.
- הכנת תא lysates
- Resuspend כדורי תא ב 1 מ"ל של פירוק מאגר (50 מ מ טריס pH 7.4, 500 מ מ NaCl, 0.4% מרחביות, 5 מ מ EDTA, 1 מ"מ DTT, 1 x מעכב פרוטאז מלאה)-מעבר RT. התאים 10 – 20 פעמים (חמש עד עשר משיכות) דרך מחט 25 גרם.
- Sonicate את הדגימות עם מכשיר sonification.
הערה: עם המכשיר sonification המצוין בטבלה של חומרים, התוכנית הבאה יכול לשמש: ארבעה מחזורים בעוצמה גבוהה, 30 לשנייה בכל מחזור התקררות (4 ° C) המים הרותחים. התקן sonification אחר מתאים אך פרמטרים אולי צריכים להיות מותאמים בהתאם. - להוסיף טריטון X-100 התאושש sonicated lysate להגיע ריכוז סופי של 2% (בדרך כלל, להוסיף 100 μL של 20% טריטון X-100 כדי 900 μL sonicated תא lysate), ולאחר מכן 2.3 מ ל 50 מ מ. טריס, pH = 7.4 לכל 1 מ ל lysate כדי להתאים את הריכוז NaCl עד 150 מ מ לפני b inding אל streptavidin-בשילוב חרוזים.
הערה ביקורתית: ריכוז המלח גבוה לעיתים קרובות לגרום הרבה פחות יעילה מחייב החרוזים. - להפיץ את lysates מנוכי עונתיות 1.5 mL צינורות (ca. מ 1.1 ל שלושה צינורות) ו centrifuge אותם ב- 16,000 x גרם, 10 דקות, 4 מעלות צלזיוס.
- להעביר את supernatants (ca. 3.2 מ"ל) שפופרת 15 mL, ו שמור 50 – 100 μL כחומר קלט.
- למדוד את ריכוז כל דגימה עם שיטת ברדפורד ומשתמשים שווה ערך של 3-3.5 מ"ג חלבון התוכן הנפתח streptavidin.
- Streptavidin הנפתח
הערה: סקירה כללית של ההליך הנפתח מוצג על איור 4. . זה כמעט זהה הפרוטוקול המקורי פורסם על-ידי רו ועמיתיו2. בפעם הראשונה הנפתח מבוצע, דגימות של מסיר כתמים זרימה דרך 1 – 4 עשוי לשים בצד לניתוח תספיג להבטיח ההליך פעל כהלכה (איור 5).- עבור כל תנאי, להעביר μL 200 של השעיה מצמידים streptavidin beads מגנטי צינור 1.5 מ. מקום הצינורות על מתלה מגנטי, המתן עד החרוזים לדבוק הצד של הצינורות (ca. 1 דקות) ולהסיר את מאגר אחסון.
- לשטוף את החרוזים על ידי ערבוב בעדינות עם 1 מ"ל של מאגר equilibration (50 מ מ טריס pH 7-4, 150 מ מ NaCl, 0.05% טריטון X-100 ו- 1 מ מ DTT).
- מוקד את החרוזים equilibrated בסכום השווה ב המספר הדרוש של צינורות (בדרך כלל כאשר החל 3-3.5 מ"ג חלבון התוכן, ש-lysates של כל תנאי ניתן שתישלח. צינורות 1.5 מ ל 2-4), המקום בחזרה על המדף מגנטי.
- הסר את המאגר equilibration, resuspend כל ערכה של חרוזים עם כמויות שוות של lysates תא התואם מהשלב 4.4.7. דגירה בין לילה ב 4 ° C על גלגל מסתובב.
- ביום הבא, במקום הצינורות 1.5 mL על מתלה מגנטי, המתן עד החרוזים לדבוק הצד של הצינורות ולהעביר את supernatants צינור 15 מ"ל כאנטי לזרום דרך.
הערה: מעכשיו, כל הצעדים מבוצעים בטמפרטורת החדר אלא אם צוין אחרת. - Resuspend את החרוזים כל צינור עם 200 μL שטיפת מאגר 1 (2% מרחביות במים), ולשלב את כל ערכה של חרוזים resuspended התואם תנאי אחד 1.5 mL צינורות.
- לשטוף את החרוזים פעמיים במשך 8 דקות על גלגל סיבוב עם 1 מ"ל של שטיפת מאגר 1.
- לשטוף את החרוזים פעמיים במשך 8 דקות על גלגל סיבוב עם 1 מ"ל של שטיפת מאגר 2 (50 מ מ HEPES pH 7.4, 1 מ"מ EDTA, 500 מ מ NaCl, 1% טריטון X-100 0.1% נה-deoxycholate).
- לשטוף את החרוזים פעמיים במשך 8 דקות על גלגל סיבוב עם 1 מ"ל של שטיפת מאגר 3 (10 מ מ טריס pH 8, 250 מ מ LiCl, 1 מ"מ EDTA, 0.5% NP-40 0.5% נה-deoxycholate).
- לשטוף את החרוזים פעמיים במשך 8 דקות על גלגל סיבוב עם 1 מ"ל של שטיפת מאגר 4 (50 מ מ טריס pH 7.4, 50 מ מ NaCl, 0.1% NP-40).
- כדי לוודא שהמאגר שטיפת מוסר לחלוטין לאחר האחרון לשטוף צעד, להסיר את מרבית תגובת שיקוע ולאחר מכן ספין למטה הדגימות. לשים אותם בחזרה על המדף מגנטי, המתן עד החרוזים לדבוק הצד של הצינורות ולאחר מכן להסיר את המאגר הנותרים.
- להוסיף 30 μL • תנאי מאגר (10 מ מ טריס pH 7.4, מרחביות 2%, 5% β-mercaptoethanol ו- 2 מ מ ביוטין) את החרוזים. דגירה ב 98 ° C למשך 15 דקות ולאחר מכן להסיר לאלתר את החרוזים על מתלה מגנטי.
- להעביר את הדגימה eluted צינור טריים ו החנות ב-20 ° C עד עיבוד נוסף.
- מרחביות-דף החיוורים ומכתים המערבי
הערה: לפני ניתוח ספקטרומטר מסה, מומלץ להעריך את ההצלחה של biotinylation, הנפתח על ידי עמודים מרחביות תספיג חלבון. אם אין דפוס biotinylation נצפית ניתן לחשוב כי dox או ביוטין לא נוסף למדיום או שאחד הפתרונות מניות שני נמצא בסיכון.- עבור כל מדגם קלט, הכן מדגם דף על ידי ערבוב כמויות שוות של חלבון לדוגמה עם נפח מתאים של מאגר מרחביות העמסה x 3 סך של 28 μL. הכן דוגמאות דף על ידי ערבוב 5 µL של כל מדגם • תנאי עם µL 2.5 מאגר מרחביות העמסה x 3.
- לטעון את הדגימות (µL 25/טוב על תשומות, 7 µL טוב עבור דגימות • תנאי) על ג'ל מרחביות-לזיהוי. המשך אלקטרופורזה ומערביות סופג כמתואר בסעיף 3.4.
- אם מבצע את הנפתח בפעם הראשונה, להכין דף הדוגמאות עבור כל שלב של טלסין (pulldown) (קלט, לזרום, • 1 – 4, תנאי לשטוף) על ידי ערבוב 20 µL של כל דגימה (קלט, זרימה דרך, שטיפת 1 – 4) עם µL 10 של 3 x מאגר מרחביות העמסה או 5 µL (• תנאי) עם 2.5 µL של 3 x S DS-טעינת מאגר והמשך כמו על שלב 4.6.4 (איור 5).
- מרחביות-דף לניתוח MS
הערה: כדי לצמצם אפשרות של זיהום קרטין, ג'לים precast ומאגר טעינה הדגימה מסחרי עשוי לשמש.- להוסיף μL 6.25 מאגר מדגם x 4 μL 18.75 של כל מדגם • תנאי ולהפעיל את הדוגמאות על 4 – 20% precast מרחביות-ג'ל עד הם נודדים 2 – 3 ס מ לתוך הג'ל.
- מכתים את הג'ל עם Coomassie colloidal G250 כחול מבריק מכתים13 בצלוחית 15 ס מ.
- השתמש באזמל נקי בלו המסלולים שלם עבור כל דגימה, למעט הלהקה streptavidin (ריצה ב- ca. 17 kDa, איור 6), ולהעביר הלהקות נכרת ל 1.5 mL צינורות.
- לשלוח דגימות אלה מתקן פרוטאומיקס לצורך ניתוח נוסף.
הערה: ניתן לראות תוצאות MS טיפוסי בהפניה 9 (איור 3 ו איור 7, וטבלאות משלים). כל קבצי הנתונים זמינים על המאגר גאווה (ההצטרפות מספר PXD005005).
תוצאות
כדי להמחיש כמה שיטה זו פועלת, מסגרות קריאה פתוחה (ORFs) של החלבונים Ago2, TNRC6C, לטעמים (הכל מעורב הגן בתיווך miRNA להשתיק לשביל) היו משובטים ב split-BioID פלסמידים. Ago2 ידוע כדי לקיים אינטראקציה עם TNRC6C בתוך miRNA-induced שתיקה קומפלקס (miRISC) represses תרגום ולעורר דעיכה של היעד mRNAs14. לפני כדי להרכיב את miRISC, Ago2 אינטראקציה עם לטעמים, האנזים שמייצר miRNAs בוגרת, בתוך מתחם שבו הוא עשוי לקבל טעון עם miRNA15. ומכאן הפיצול-BioID הוחל על הזוג Ago2/לטעמים או הזוג Ago2/TNRC6C. עבור כל זוג של חלבונים שנבדקו, Ago2 היה גם התמזגו NBirA * או CBirA * שימוש שלנו פלסמידים פיצול-BioID (איור 2), לטעמים, TNRC6C המקביל cognate בירה * קטע. בנוסף, כל חלבון הייתה דבוקה CBirA *, יחד עם NBirA *-GFP פיוז'ן כפקד שלילי. התוצאה בדיקות ארבע איטרציות עבור כל זוג של נבדק חלבון (טבלה 1).
כדי לבדוק אם פיצול-BioID מופעל על האינטראקציה של הזוג של חלבונים שנבדקו, עקבנו ערכת מתואר באיור1. פלסמידים היו transfected transiently קו ט-מערכת תואם התא הלה. הביטוי של החלבונים פיוז'ן הושרה עם דוקסיציקלין (dox), biotinylation היה מגורה על ידי הוספת ביוטין עודף מדיום הגידול. לאחר זמן הדגירה 20 h עם dox, ביוטין, התאים היו lysed, נותחו על ידי המערב סופג באמצעות streptavidin מצומדת לזהות חלבונים biotinylated. בתרבית של תאים, שתי להקות העיקריים הם בדרך כלל שזיהה את streptavidin מצומדת במדגם untransfected (איור 3, כוכבים) ומתייחסים אליהם endogenously biotinylated חלבונים (ככל הנראה מיטוכונדריאלי carboxylases). לצד להקות אלו קיימים כל דוגמאות והוא יכול לשמש בנוחות כפקדי הטעינה פנימי, לפיכך, הגילוי של חלבון משק הבית כדי לשלוט הטעינה של כמויות חלבון שווה הוא מיותר. אופייני עבור ניסוי BioID/פיצול-BioID, הלהקות הגדולות נוספים, זה יכול להיות שנצפו הם החלבונים פיוז'ן שיש העצמי-biotinylated. גם אם אין חלבונים biotinylated אחרות נתפסת, גילוי biotinylation של החלבונים פיוז'ן בשלב זה כבר מציין שני החלבונים שנבדקו תקשרתי בתאים. בניסוי המתואר באיור3, ברור כי בעל NBirA של *-חלבון כימרי Ago2 יחד עם CBirA * fusions TNRC6C או מקצץ יעיל יותר מאשר צירופי מול איזה CBirA *-Ago2 מזווג כדי NBirA * fusions של השניים האחרים חלבונים (איור 3, לוח העליון, להשוות עוצמות בשבילים 2-3 עד מסלולים 6-7). יתר על כן, ההפעלה הייתה ספציפית כמו אף אחד CBirA * fusions יכול להפעיל את NBirA *-GFP שליטה חלבון כימרי לרמות ניכר (איור 3, להשוות בין סמטאות 1, 4-5 ליין 8 שמתאים לתאים untransfected). מאז פלסמידים שלנו, NBirA * יש תג myc ו CBirA * יש תג דגל (איור 2), ניתן לנתח את רמות הביטוי של כל חלבון כימרי עם נוגדנים נגד אלה שני תגים (איור 3, החלונית התחתונה).
כאשר נוצרת אינטראקציה-induced biotinylation, הניסוי ניתן לשנות, החלבונים biotinylated מבודדת על חרוזים מצמידים streptavidin כמצוין בפסקה 4 של הפרוטוקול (איור 4). בעת ביצוע הבידוד בפעם הראשונה, כל השלבים של הטיהור עשוי להיות מנותח על ידי המערב סופג (איור 5). בדרך כלל, מחייב החרוזים צריך להיות כמותיים כמעט, כמעט אין דליפה דרך להתייחס בשוטף. ראש המנזר עיבוד הדגימות עבור ספקטרומטר מסה, אנו ממליצים פועל תספיג חלבון כדי להבטיח המושרה-biotinylation עבד כצפוי החלבונים פיוז'ן באו לידי ביטוי. העדר ביטוי החלבונים היתוך הוא עקב יעילות תרביות תאים עניים או אינדוקציה dox פגום או. אם החלבונים פיוז'ן באו לידי ביטוי, אך biotinylation לא נצפית, בדוק אם עודף ביוטין (50 μM) נוספה למעשה המדיום ביוטין מניות זה עדיין פעיל. כאשר החומר eluted ניתוח על ג'ל שהוכתמו Coomassie חלבון (איור 6), בדרך כלל, הלהקה הכי חזק להיות נצפתה בנכסיה 17 kDa, מקביל monomeric streptavidin. גם עלולים להיות שנצפו להקות התואם את החלבונים biotinylated אנדוגני וחלבונים פיוז'ן. . אנחנו נכנסים בדרך כלל האזור של השביל מדגם מעל הלהקה streptavidin עד ההעמסה טוב (איור 6). הלהקה נכרת יכול להיות מאוחסן צינור 1.5 מ ל ונשלח למתקן ספקטרומטר מסה. לחלופין, חלבונים מאוגד יכול להיות גם טריפסין-מעוכלים על החרוזים מצמידים streptavidin ויוצרים פפטידים מתעכל eluted את העמודה. אנו משתמשים באופן שגרתי את MaxQuant תוכנה16 (שימוש בעיקר ברירת המחדל בפרמטרים, תוך הוספת ליזין biotinylation post-translational שינוי אפשרי, ראה חומר עיון בנושא 9 לפרטים נוספים, לתוצאות MS טיפוסי) כדי לנתח את הנתונים הגולמיים של MS ו פרסאוס סוויטה17 עבור ניתוח סטטיסטי עוקבות, שניהם תוכנה חופשית. דוגמאות מופעלים בדרך כלל משכפל ביולוגי שלוש. באמצעות כימות ללא תווית, ניתן לזהות באופן ספציפי מועשר חלבונים על תנאי הבקרה. כדי לבצע סינון עבור endogenously biotinylated חלבונים, חלבונים המסומנות הלא ספציפית על ידי האנזים בירה *, אנו רואים רק חלבונים כי הם מועשרים באופן משמעותי על להיטים מתוך שישה datasets שנוצר עם שישה חלבונים לא קשורים. בנוסף, אנו רואים רק כניסות מועשרים מעל dataset פיצול-BioID שבו פיוז'ן החלבונים NBirA * הוחלפו על-ידי NBirA *-GFP. אסטרטגיות ניתוח נתונים אחרים הוצעו בעיקר באמצעות איזוטופ יציב תיוג עם חומצות אמינו בתרבות תא (SILAC) עבור פרוטאומיקס כמותיים18. בנוסף, אסטרטגיות שונות תוארו של בידוד ישיר של פפטידים biotinylated באמצעות משתנה streptavidin עם זיקה חלשה ביוטין18, תנאים מיוחדים • תנאי שימוש בממסים אורגניים19 או ספציפיים ביוטין נוגדנים20,21. בזמן לא בהכרח מובילים לגילוי של חלבונים נוספים, זיהוי אתרים biotinylation להוסיף יותר ביטחון לגבי יחודיות של הלהיטים, היא שימושית בעת מיעון את הטופולוגיה של אינטראקציה.

איור 1: סקירה של ההליך פיצול-BioID. חלבון 1 אינטראקציה עם חלבון 2 חלק א מורכבים, או עם חלבון 3 חלק ב' מורכב לחקור באופן ספציפי את ההרכב של מורכבות A, ניתן להחיל פיצול-BioID חלבונים 1 ו- 2. הצילום של ספקטרומטר מסה תחת רישיון יצירתי של מלאי ייחוס-שיתוף כאחד 3.0 לא מותאם, שהורד מן https://commons.wikimedia.org עם שם קובץ של ThermoScientificOrbitrapElite.JPG. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2: ביטוי קלטות של פלסמידים פיצול-BioID. אנו מספקים פלסמידים ארבע כדי לאפשר בדיקה כל השילובים של NBirA *, CBirA * פיוז'ן חלבונים. פלסמידים ומפות מלא הינם זמינים addgene.org תחת המספרים המצוינים. פלסמידים יש אלמנט מגיב טט (7 x tetO), צריך לשמש קו תא התואמת המערכת ביטוי ט. גם שים לב ב פלסמידים כל ORFs של FKBP ואת FRB הם התמזגו כדי NBirA *, CBirA * קטעים בהתאמה. אלה שני חלבונים אינטראקציה רק בנוכחות rapamycin, ומכאן פלסמידים יכול לשמש כדי לבדוק במהירות את המערכת נוכחות או העדר הזה כימי9. האתרים הגבלת המצוין הם ייחודיים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3: כתם המערבי הטיפוסי עבור ניסוי פיצול-BioID. פאנל עליון: זיהוי של חלבונים biotinylated עם streptavidin fluorescently עם תוויות. החלונית התחתונה: זיהוי של החלבונים פיוז'ן עם נוגדנים אנטי-Myc ודגל אנטי. שני זוגות של חלבונים נבדקו: Ago2/TNRC6C ו- Ago2/לטעמים. בנתיבים 2 & 3, Ago2 היה מצורף למקטע CBirA *. בנתיבים 6 & 7, Ago2 היה מצורף למקטע NBirA *. אין אות משמעותי נצפתה כאשר כל אחד החלבונים 3 משולב עם NBirA *-GFP (מסלולים 1, 4-5). הכוכבים מציינים הלהקות המתאים endogenously חלבונים biotinylated יכול לשמש כפקדי הטעינה פנימי. נתון זה ממאמרו של איור 5B של. Schopp et al. 9 תחת רישיון יצירתי של מלאי ייחוס 4.0 הבינלאומי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 4: סקירה של ההליך טלסין (pulldown) streptavidin. מתוארים שלבים עיקריים בידוד של חלבונים biotinylated לניתוח ספקטרומטר מסה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 5: כתם המערבי הטיפוסי עבור ניסוי טלסין (pulldown) streptavidin. שווה כרכים של כל מדגם המצוין נטענו על ג'ל מרחביות-לזיהוי. בעקבות סופג המערבי, biotinylated חלבונים אותרו מצמידים HRP streptavidin. להקות המתאים NBirA *-TNRC6C ו- CBirA *-Ago2 מצוינים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 6: חלבון שהוכתמו טיפוסי Coomassie ג'ל לניתוח ספקטרומטר מסה. המדגם eluted של חרוזים מצמידים streptavidin היה טעון על ג'ל חלבון precast, לרוץ עד המדגם להעביר 2-3 ס מ. הלהקה הגדולים לראותם 17 kDa הוא streptavidin. האזור ישירות מעל הלהקה הזאת היא טוחנות ושלח למתקן ספקטרומטר מסה. להקות המתאים NBirA *-TNRC6C ו- CBirA *-Ago2 מצוינים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| תרביות תאים מדגם | תנאי נבדק | ||
| 1 | NBirA *-protein1 / CBirA *-protein2 | ||
| 2 | CBirA *-protein1 / NBirA *-protein2 | ||
| 3 | NBirA *-GFP / CBirA *-protein1 | ||
| 4 | NBirA *-GFP / CBirA *-protein2 | ||
| 5 | אין תקנים | ||
טבלה 1: בדרך כלל נבדק בתנאים בשעת החלת פיצול-BioID על שני חלבונים.
| רצף פריימר | רצף |
| קלטת 1 הפוך פריימר (CBirA * fusion) | TATACTTTCTAGAGAATAGGAAC |
| קלטת 2 פריימר הפוכה (NBirA * fusion) | GTGGTTTGTCCAAACTCATC |
טבלה 2: קביעת רצף צבעי יסוד פלסמידים פיצול-BioID.
Discussion
ההליך מחולקת לרמות מתאר איך לשכפל את הגנים של עניין לתוך הפיצול-BioID פלסמידים, כיצד לבדוק את האינטראקציה-induced biotinylation וכיצד לבודד חלבונים biotinylated לניתוח ספקטרומטר מסה. נתאר כאן פרוצדורה המבוססת על תקנים ארעית. בעוד הביטוי של החלבונים פיוז'ן ניתן לכוונן לפי כמות dox הוסיף למדיום, תרביות תאים ארעי עלול להוביל ביטוי חלבון שאינו הומוגני עם כמה תאים בגסות overexpress את החלבונים פיוז'ן בהשוואה למצבם אנדוגני עמיתיהם. זה עלול לגרום עיוותים של interactomes המתאים וכדי PPI אינן משקפות נאמנה את אינטראקציות המערבות את החלבונים אנדוגני. רצוי לכן בדרך כלל לבנות שורות תאים יציב לאחר פיצול-BioID הוקם באמצעות מערכת ארעית. פלסמידים עולים בקנה אחד עם מערכת רקומבינציה בתיווך Flp ולמקם בשני הגנים של עניין תחת ברגולציה של אותו רכיב ט מגיב. אם יש צורך, בעת שימוש עם תאים בתרבית של תואם, הם מאפשרים יצירה קלה של שורות תאים inducible יציב. לדוגמה, אנו משתמשים הקו הלה-EM2-11 המבטאת activator מופעל טטרציקלין שעתוק rtTA וגנומית ייחודי targetable לוקוס שממנו ניתן ביטוי גנים בתיווך טטרציקלין בחוזקה מוסדר12. באמצעות תאים בטור הזה, בתיווך Flp רקומבינציה, שורות תאים יציב המכילים רק עותק אחד של transgene ניתן לקבל בתוך שבועיים-שלושה שבועות. לחלופין, אחד יכול גם להשתמש טכניקות עריכה הנוכחי של הגנום להציג שברי בירה * ב לוקוסים גנומית מקורית של הגנים של עניין.
כמו כל assay המסתמך על תיוג חלבון, צריך לשקול אם החלבונים פיוז'ן וכתוצאה מכך פועלים. נתונים זמינים ב אשר תויגו החלבונים עניין (לדוגמה עם GFP הדמיה מחקרים) והן מבחינה תפקודית נבדק שימושיים כדי להחליט אם השברים בירה * צריך להיות משובטים במעלה או במורד הגנים של עניין. אם אין נתונים זמינים, אחד צריך לבדוק N-סופני או C-סופני מתויג חלבונים ב- assay פונקציונלי. לדוגמה, הפעילות של החלבונים פיוז'ן יכול להיבדק בקו התא בו החלבון אנדוגני יש כבר הפילו-אאוט, לעומת המצב פראי סוג. אם החלבונים עניין לסבול שתי תגיות N - ו C-מסוף, שניהם צריך להיבדק. אכן, בניסויים BioID, הכיוון של החלבון פיוז'ן יכול להשפיע את היעילות של תיוג22. בנוסף, הבחנו כי בעת החלת פיצול-BioID זוג של חלבונים, אשר של שני החלבונים מצורף לשם גם NBirA * או CBirA * קטע גם השפעה את היעילות של תיוג9. ב פלסמידים פיצול-BioID, חומצת אמינו 16 ארוך גליצין/סרין linkers עשירה צימוד החלבונים עניין כדי השברים בירה * היו שנלקחו PCA עוד23 , עבדה בשבילנו כל החלבונים שמעצבת בדקנו עד כה. עם זאת, ניתן לשקול כמה זוגות חלבון יכול לעבוד טוב יותר עם linkers ארוכה או קצרה יותר. ראוי לציין הסופי, וזמינותו אחר תוארה על ידי קבוצת Bollen24. ב הזה assay, בירה * מפוצל באתר אחר (E140/Q141) משלנו (E256/G257). לנו יש שני פיצול-BioID טעמים-לצד נבדק ונמצא כי E256/G257, שמתואר פרוטוקול זה, מוביל חזק הפעלה מחדש כאשר מצמידים שני חלבונים שמעצבת9.
חסרון אחד כללי של שיטה זו הוא המהירות האיטית של תיוג. בדרך כלל, זמן הדגירה 6 עד 24 h עם ביוטין יש צורך להשיג ניכר biotinylation6, מסלק את השימוש בטכניקה זו ללמוד שיפוץ דינמי של מתחמי חלבון. בזמן הזה assay כתובות חלקית זו אזהרה כמו זה מופעל רק כאשר שני חלבונים אינטראקציה, מהירות איטית של תיוג למנוע השימוש ללמוד מענה לתהליכים מאוד דינמי או לנתח חלבונים קצרת ימים. Peroxidase מהונדסים APEX2 ידוע לקדם תיוג יעיל של חלבונים proximal תוך מינימום 13. PCA בהתבסס על APEX2 ובכך אולי כתובת המגבלות של מהירות תיוג איטי מבחני נגזר BioID. מחקר הוכחה של עקרון תיאר כזה של פיצול-APEX2 assay25. עם זאת, למרות חלבון homodimerizing היה בהצלחה biotinylated, אם וזמינותו יכול לשמש גם תווית ולזהות חלבונים להרכיב סביב זוג של חלבונים שמעצבת נשאר להדגים. ממש לאחרונה, אבולוציה מכוונת שימש ליצירת TurboID, miniTurbo, שתי גרסאות של בירה * עם פעילות משופרת לאפשר חלונות זמן תיוג הרבה יותר קצר, עד 10 דקות26. התאמת פיצול-BioID אלה גרסאות חדשות נוספות תרחיב את השימוש בטכניקה זו לשדה רחבה של יישומים.
Disclosures
המחברים אין לחשוף.
Acknowledgements
עבודה זו הוקם במימון המועצה הגרמנית למחקר (DFG) דרך יוזמה מצוינות הגרמני (CellNetworks DFG-סיפורה ה 81), מימון חלקי על-ידי מרכז מחקר משותף SFB638.
Materials
| Name | Company | Catalog Number | Comments |
| Acetic acid (glacial) | VWR | 20104.298 | To make TAE buffer |
| Agarose | Sigma | A9539 | Take TAE-agarose gels for DNA analysis and extraction |
| Ammonia solution 25% NH3 | Bernd Kraft | 6012 | To dissolve biotin |
| Ampicillin | Sigma | A9518 | To select transformed bacteria |
| Bioruptor plus sonification device | Diagenode | B01020001 | Other sonification devices are also ok |
| Biotin | Sigma | B4639 | To be added to the growth medium to stimulate efficient biotinylation |
| Bovine serum albumin fraction V | Carl Roth | 8076 | Used in Western blot buffers and a protein standard in Bradford assays |
| Bradford Ultra reagent | Expedeon | BFU05L | Any other method/kit for protein determination is fine, this particular reagent is more tolerant to detergent than other Bradford reagents |
| Cell scrappers | TPP | 99002 | Any other model is also fine |
| ClaI | New England Biolabs | R0197 | Restriction enzyme for cloning into the split-BioID plasmids (NBirA* fusion) |
| DMEM medium | Sigma | D6046 | If using another cell line, use the corresponding optimal growth medium |
| DNA miniprep kit | Sigma | PLN350 | Any other kit is also fine |
| Doxycycline | Applichem | A2951 | Dox is light sensitive |
| DTT | Applichem | A2948 | Make 1M stock solution, store at -20 °C and always use fresh |
| DyLight 680-conjugated streptavidin | Thermo scientific | 21848 | to use with a LiCor Western blot scanning device |
| Dynabeads MyOne Streptavidin C1 | Invitrogen | 65002 | The C1 beads are not BSA coated which is preferable for downstream MS applications (no leakthrough of BSA in the final elution) |
| EDTA | Applichem | A5097 | Make a 500 mM stock, adjust pH to 8 while dissolving the EDTA powder |
| Ethanol | Sigma | 32205 | Make a 70% stock solution in which Doxycycline can be dissolved at 10 mg.mL-1 |
| Fastgene Gel/PCR DNA Extraction Kit | Nippon Genetics | FG-91302 | Any other kit is also fine |
| HCl 37% | Merck | 1.00317.1000 | To adjust pH of biotin stock solution |
| HEPES | Carl Roth | 6763 | Make a 500 mM stock solution, adjust the pH to 7.4 |
| Immobilon-FL PVDF membrane, 0.45 µm | Millipore | IPFL00010 | This membrane shows minimal autofluorescence when used with a LiCor Western blot scanning device |
| LiCl | Grüssing GmbH | 12083 | Make a 5M stock solution |
| Odyssey CLx imaging system | LI-COR | N/A | To scan Western blot membrane decorated with fluorophore-labeled antibody |
| Linear polyethylenimine (PEI) | Polysciences | 23966-2 | Any other transfection reagent is also fine |
| Milk powder | Carl Roth | T145 | To block Western blot membranes |
| MluI-HF | New England Biolabs | R3198 | Restriction enzyme for cloning into the split-BioID plasmids (NBirA* fusion) |
| Na-deoxycholate | Sigma | 30970 | Make a 10% (w/v) stock solution |
| NaCl | Sigma | 31434 | Make a 5M stock solution |
| NP-40 (Nonidet P40 substitute) | Sigma | 74385 | Make a 20% (v/v) stock solution |
| PacI | New England Biolabs | R0547 | Restriction enzyme for cloning into the split-BioID plasmids (CBirA* fusion) |
| Phosphate buffer saline (PBS) | Sigma | 806552 | To wash cells before scrapping |
| PmeI | New England Biolabs | R0560 | Restriction enzyme for cloning into the split-BioID plasmids (CBirA* fusion) |
| Protease inhibitor cocktail | Roche | 4693132001 | Added to the lysis buffer to prevent protein degradation |
| pSF3-Flag-CBir-FRB_Myc-NBir-FKBP | Addgene | 90003 | Split-BioID plasmid, mediates the co-expression of NBirA*-FKBP and CBirA*-FRB, FKBP and FRB can be replaced by two other ORFs |
| pSF3-Flag-CBir-FRB_FKBP-Myc-Nbir | Addgene | 90004 | Split-BioID plasmid, mediates the co-expression of FKBP-NBirA* and CBirA*-FRB, FKBP and FRB can be replaced by two other ORFs |
| pSF3-Flag-FRB-Cbir_Myc-NBir-FKBP | Addgene | 90008 | Split-BioID plasmid, mediates the co-expression of NBirA*-FKBP and FRB-CBirA*, FKBP and FRB can be replaced by two other ORFs |
| pSF3-Flag-FRB-Cbir_FKBP-Myc-Nbir | Addgene | 90009 | Split-BioID plasmid, mediates the co-expression of FKBP-NBirA* and FRB-CBirA*, FKBP and FRB can be replaced by two other ORFs |
| Q5 High-Fidelity PCR kit | New England Biolabs | E0555S | To amplify the ORF coding for the proteins to be tested. Any other thermostable DNA polymerase is fine. |
| Quick ligation kit | New England Biolabs | M2200S | To ligate DNA fragments into the split-BioID plasmids, any other DNA ligation system is fine. |
| RunBlue 4-20% SDS precast gels | Expedeon | BCG42012 | To use when running samples for MS analysis |
| RunBlue LDS Sample Buffer | Expedeon | NXB31010 | Running buffer for the RunBlue precast gels |
| SDS | Sigma | 5030 | Comes as a 20% stock solution |
| tet-free serum | Biowest | S181T | we use tet-free serum to minimize basal expression of the fusion proteins |
| Trans-Blot Turbo Transfer system | Bio-Rad | 1704150 | High speed Western blotting transfer system, any other transfer system is also fine |
| Tris | Carl Roth | 4855 | Make 1M stock solutions with adequate pH (7.4 and 8) |
| Triton X-100 | Applichem | A4975 | Make a 20% (v/v) stock solution |
| Tween-20 | Carl Roth | 9127 | Used in Western blot buffers, Tween 20 leads to high background fluorescence and should be omitted in the blocking and last wash step |
References
- Scott, D. E., Bayly, A. R., Abell, C., Skidmore, J. Small molecules, big targets: drug discovery faces the protein-protein interaction challenge. Nat Rev Drug Discov. 15 (8), 533-550 (2016).
- Roux, K. J., Kim, D. I., Raida, M., Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J Cell Biol. 196 (6), 801-810 (2012).
- Lam, S. S., et al. Directed evolution of APEX2 for electron microscopy and proximity labeling. Nat Methods. 12 (1), 51-54 (2015).
- Rhee, H. W., et al. Proteomic mapping of mitochondria in living cells via spatially restricted enzymatic tagging. Science. 339 (6125), 1328-1331 (2013).
- Kim, D. I., et al. Probing nuclear pore complex architecture with proximity-dependent biotinylation. P Natl Acad Sci USA. 111 (24), E2453-E2461 (2014).
- Kim, D. I., et al. An improved smaller biotin ligase for BioID proximity labeling. Mol Biol Cell. 27 (8), 1188-1196 (2016).
- Lambert, J. P., Tucholska, M., Go, C., Knight, J. D., Gingras, A. C. Proximity biotinylation and affinity purification are complementary approaches for the interactome mapping of chromatin-associated protein complexes. J Proteomics. 118, 81-94 (2015).
- Morriswood, B., et al. Novel bilobe components in Trypanosoma brucei identified using proximity-dependent biotinylation. Eukaryot Cell. 12 (2), 356-367 (2013).
- Schopp, I. M., et al. Split-BioID a conditional proteomics approach to monitor the composition of spatiotemporally defined protein complexes. Nat Commun. 8, (2017).
- Béthune, J., Artus-Revel, C. G., Filipowicz, W. Kinetic analysis reveals successive steps leading to miRNA-mediated silencing in mammalian cells. EMBO Rep. 13 (8), 716-723 (2012).
- Inoue, H., Nojima, H., Okayama, H. High efficiency transformation of Escherichia coli with plasmids. Gene. 96 (1), 23-28 (1990).
- Weidenfeld, I., et al. Inducible expression of coding and inhibitory RNAs from retargetable genomic loci. Nucleic Acids Res. 37 (7), e50(2009).
- Dyballa, N., Metzger, S. Fast and sensitive colloidal coomassie G-250 staining for proteins in polyacrylamide gels. J Vis Exp. (30), (2009).
- Jonas, S., Izaurralde, E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat Rev Genet. 16 (7), 421-433 (2015).
- MacRae, I. J., Ma, E., Zhou, M., Robinson, C. V., Doudna, J. A. In vitro reconstitution of the human RISC-loading complex. P Natl Acad Sci USA. 105 (2), 512-517 (2008).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol. 26 (12), 1367-1372 (2008).
- Tyanova, S., et al. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat Methods. 13 (9), 731-740 (2016).
- Opitz, N., et al. Capturing the Asc1p/Receptor for Activated C Kinase 1 (RACK1) Microenvironment at the Head Region of the 40S Ribosome with Quantitative BioID in Yeast. Mol Cell Proteomics. 16 (12), 2199-2218 (2017).
- Mackmull, M. T., et al. Landscape of nuclear transport receptor cargo specificity. Mol Syst Biol. 13 (12), 962(2017).
- Kim, D. I., et al. BioSITe: A Method for Direct Detection and Quantitation of Site-Specific Biotinylation. J Proteome Res. , (2017).
- Udeshi, N. D., et al. Antibodies to biotin enable large-scale detection of biotinylation sites on proteins. Nat Methods. 14 (12), 1167-1170 (2017).
- Chapat, C., et al. Cap-binding protein 4EHP effects translation silencing by microRNAs. P Natl Acad Sci USA. 114 (21), 5425-5430 (2017).
- Luker, K. E., et al. Kinetics of regulated protein-protein interactions revealed with firefly luciferase complementation imaging in cells and living animals. P Natl Acad Sci USA. 101 (33), 12288-12293 (2004).
- De Munter, S., et al. Split-BioID: a proximity biotinylation assay for dimerization-dependent protein interactions. FEBS Lett. 591 (2), 415-424 (2017).
- Xue, M., et al. Optimizing the fragment complementation of APEX2 for detection of specific protein-protein interactions in live cells. Sci Rep. 7 (1), 12039(2017).
- Branon, T. C., et al. Directed evolution of TurboID for efficient proximity labeling in living cells and organisms. bioRxiv. , (2017).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved